តើអ្វីជាភាពខុសគ្នារវាង Synthase និង Synthetase? (ការពិតត្រូវបានបង្ហាញ) - ភាពខុសគ្នាទាំងអស់។

តារាងមាតិកា
អង់ស៊ីម ligase E.C. 6 រួមមានសំយោគ និងសំយោគ។ ពួកវាចូលរួមក្នុងប្រតិកម្មសំយោគ និងជំរុញការរួមផ្សំនៃម៉ូលេគុលពីរ ក្នុងពេលដំណាលគ្នាបំបែកតំណភ្ជាប់ diphosphate នៅក្នុង ATP ឬ triphosphate ប្រៀបធៀបមួយផ្សេងទៀត។
ផ្ទុយទៅនឹងសំយោគ ដែលប្រើ ATP ជាប្រភពថាមពលកំឡុងពេលសំយោគ នៃសមាសធាតុជីវសាស្រ្ត សំយោគគឺជា ligases ដែលជំរុញការសំយោគនៃសមាសធាតុជីវសាស្រ្តដោយមិនប្រើ ATP ជាប្រភពថាមពល។
នៅក្នុងអត្ថបទនេះ អ្នកនឹងដឹងច្បាស់ពីភាពខុសគ្នារវាង Synthase និង Synthetase។
តើ Synthase ជាអ្វី?
សារធាតុសំយោគគឺជាអង់ស៊ីមដែលជំរុញដំណើរការសំយោគនៅក្នុងជីវគីមី។ សូមចងចាំថានាមត្រកូលជីវសាស្រ្តដំបូងត្រូវបានបែងចែករវាងសំយោគនិងសំយោគ។
ដោយអនុលោមតាមនិយមន័យដើម ការសំយោគប្រើប្រាស់ nucleoside triphosphates (ដូចជា ATP, GTP, CTP, TTP និង UTP) ជាប្រភពថាមពល ខណៈពេលដែលសំយោគមិនមាន។
ទោះជាយ៉ាងនេះក្តី យោងទៅតាមគណៈកម្មការចម្រុះស្តីពីឈ្មោះគីមីជីវៈ (JCBN) "ការសំយោគ" អាចត្រូវបានប្រើដើម្បីយោងទៅលើអង់ស៊ីមណាមួយដែលជំរុញការសំយោគ (ដោយមិនគិតពីថាតើវាប្រើ nucleoside triphosphates) ប៉ុន្តែ "សំយោគ ” គួរតែត្រូវបានប្រើសម្រាប់តែសំដៅទៅលើ “ligase ប៉ុណ្ណោះ។ Citrate synthase
ភាពខុសគ្នាចម្បងរវាង Synthase និង Synthetase គឺ ថា Synthetase គឺជាអំបូរនៃអង់ស៊ីមដែលអាចបង្កើតចំណងរវាងម៉ូលេគុល ចំណែក Synthase គឺជាអង់ស៊ីម។
| Synthase | Synthetase |
| កាតាលីករដំណើរការសំយោគដោយគ្មាន ATP | ទាមទារ ATP |
| មាននៅក្រោមការចាត់ថ្នាក់ vase ឬ transferase | មកក្រោមការចាត់ថ្នាក់ ligase |
| ឧ. HMG-COA synthase, ATP synthase | ឧ. Succiny1-COA synthetase, Glutamine synthetase |
Synthase vs Synthetase តារាងប្រៀបធៀប
មើលវីដេអូនេះ ដើម្បីដឹងពីការសំយោគ និងសំយោគ
សេចក្តីសន្និដ្ឋាន
- ការសំយោគមិនត្រូវការ NTPs ដើម្បីដំណើរការទេ ចាប់តាំងពីពួកគេបានជំរុញដំណើរការសំយោគដែលត្រូវការអ៊ីដ្រូលីស៊ីតនៃនុយក្លេអូស៊ីត triphosphates (មិនមែនគ្រាន់តែ ATP) នោះទេ។
- សហភាពអន្តរជាតិនៃ គណកម្មាធិការនាមត្រកូលរបស់ជីវគីមីវិទ្យាបានផ្លាស់ប្តូរនិយមន័យនៃ synthase ក្នុងទសវត្សរ៍ឆ្នាំ 1980 ដើម្បីរួមបញ្ចូលអង់ស៊ីមសំយោគទាំងអស់ ថាតើពួកគេបានប្រើប្រាស់ NTPs ឬអត់ ហើយការសំយោគបានក្លាយទៅជាមានន័យដូច ligase ។
- Ligase គឺជាអង់ស៊ីមដែលភ្ជាប់ម៉ូលេគុលតូចៗចំនួនពីរជាមួយគ្នាដោយការប្រើប្រាស់ ថាមពលពី NTP hydrolysis (ជាធម្មតាតាមរយៈ condensation មួយ។ប្រតិកម្ម)
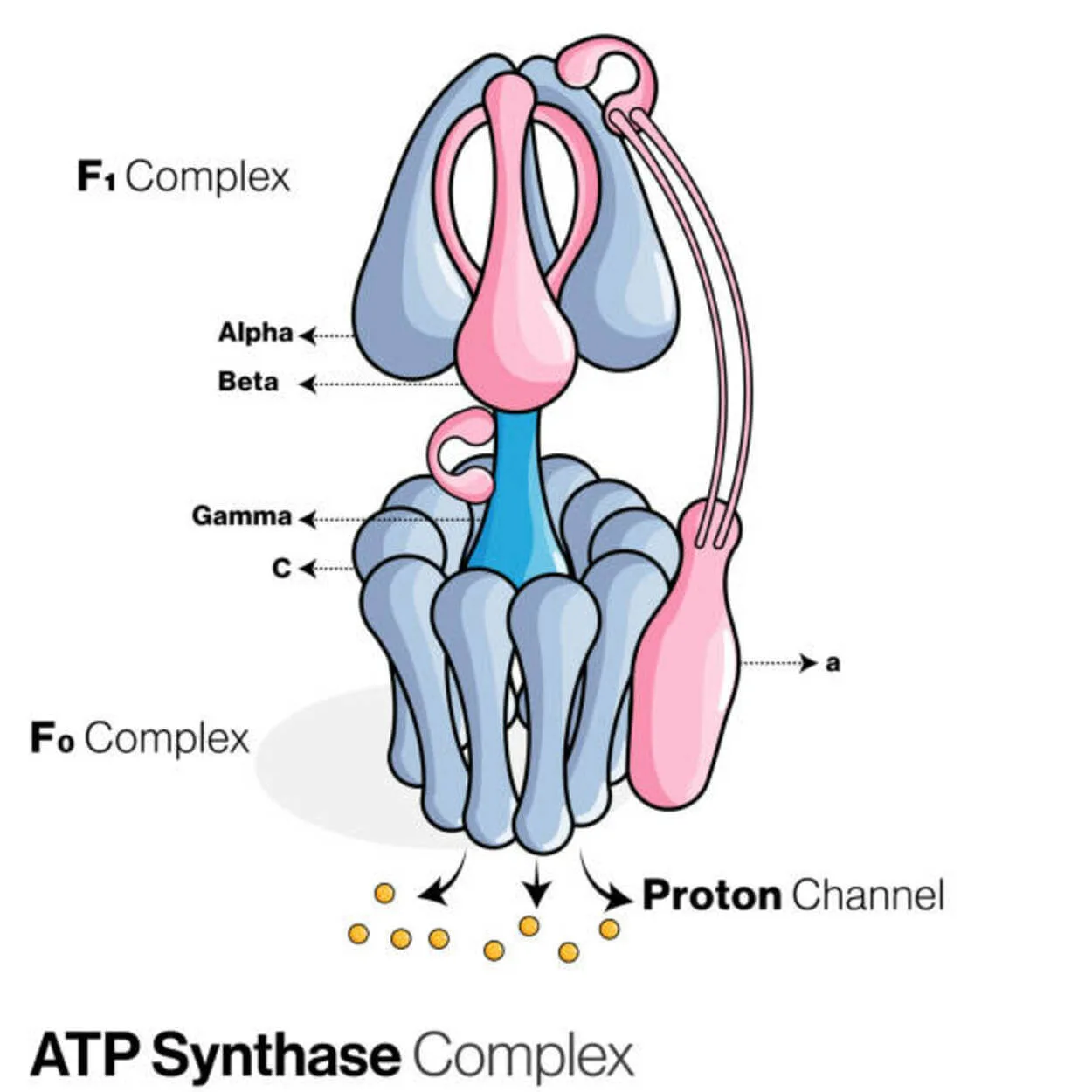
ATP Synthase
Adenosine diphosphate (ADP) និង phosphate inorganic ត្រូវបានប្រើដើម្បីបង្កើតម៉ូលេគុលផ្ទុកថាមពល adenosine triphosphate (ATP) ដោយប្រូតេអ៊ីនហៅថា ATP synthase (Pi)។
វាត្រូវបានចាត់ថ្នាក់ជា ligase ចាប់តាំងពីវាកែប្រែ ADP ដោយបង្កើតជាតំណភ្ជាប់ P-O (ចំណង phosphodiester) ។ ឧបករណ៍ម៉ូលេគុលដែលហៅថា ATP synthase។
ដោយថាមពល ការផលិត ATP ពី ADP និង Pi គឺមិនចង់បាន ហើយដំណើរការនេះជាធម្មតាទៅវិធីផ្សេង។
ជម្រាលកំហាប់ប្រូតុង (H+) ឆ្លងកាត់ភ្នាស mitochondrial ខាងក្នុងនៅក្នុង eukaryotes ឬភ្នាសប្លាស្មានៅក្នុងបាក់តេរីជំរុញឱ្យប្រតិកម្មនេះទៅមុខដោយភ្ជាប់ការសំយោគ ATP កំឡុងពេលដកដង្ហើមកោសិកាទៅជម្រាល។
នៅក្នុងរុក្ខជាតិ ATP synthase ប្រើជម្រាលប្រូតុងដែលបង្កើតឡើងនៅក្នុង thylakoid lumen ឆ្លងកាត់ភ្នាស thylakoid និងចូលទៅក្នុង chloroplast stroma ដើម្បីផលិត ATP កំឡុងពេលធ្វើរស្មីសំយោគ។
សម្រាប់ ATPase ការសំយោគ ATP eukaryotic គឺ F -ATPases ដែលមានមុខងារ "បញ្ច្រាស" ។ ប្រភេទនេះត្រូវបានពិភាក្សាជាចម្បងនៅក្នុងអត្ថបទនេះ។ អនុក្រុម FO និង F1 នៃ F-ATPase មានយន្តការបង្វិលដែលអាចឱ្យការសំយោគ ATP ។
មានប្រភេទផ្សេងគ្នានៃ Synthase
Citrate Synthase
ស្ទើរតែគ្រប់កោសិការស់ទាំងអស់មានផ្ទុកអង់ស៊ីម citrate synthase,ដែលដើរតួនាទីជាអ្នកជំរុញល្បឿនក្នុងជំហានដំបូងនៃវដ្តអាស៊ីតនៃក្រូចឆ្មា និងត្រូវបានកំណត់ E.C. 2.3.3.1 (ពីមុន 4.1.3.7)។ (ឬវដ្ត Krebs) ។
Citrate synthase មានទីតាំងនៅក្នុងម៉ាទ្រីស mitochondrial នៃកោសិកា eukaryotic ទោះបីជា DNA នុយក្លេអ៊ែរ មិនមែន DNA mitochondrial អ៊ិនកូដវាក៏ដោយ។
វាត្រូវបានបង្កើតនៅក្នុង cytoplasm ដោយ cytoplasmic ribosomes ហើយបានផ្លាស់ប្តូរជាបន្តបន្ទាប់ទៅក្នុង mitochondrial matrix។
សញ្ញាសម្គាល់អង់ស៊ីមបរិមាណធម្មតាសម្រាប់អត្ថិភាពនៃ mitochondria ដែលនៅដដែលគឺ citrate synthase។ សកម្មភាពកំពូលរបស់ Citrate synthase បង្ហាញពីចំនួន mitochondria មានវត្តមាននៅក្នុងសាច់ដុំគ្រោងឆ្អឹង។
ការហ្វឹកហ្វឺនចន្លោះពេលអាំងតង់ស៊ីតេខ្ពស់មានសក្តានុពលក្នុងការបង្កើនសកម្មភាពអតិបរិមានៃលើសពីការហ្វឹកហ្វឺនការស៊ូទ្រាំ ឬការហ្វឹកហ្វឺនចន្លោះពេលអាំងតង់ស៊ីតេខ្ពស់។
អាសេទីលកូអង់ស៊ីម A មានសំណល់អាសេតាតកាបូនពីរ និងម៉ូលេគុលនៃបួន។ -carbon oxaloacetate condenses ដើម្បីបង្កើតកាបូនប្រាំមួយ citrate ដែលត្រូវបានផលិតដោយប្រតិកម្ម condensation កាតាលីករដោយ citrate synthase។
Tryptophan Synthase
ជំហានពីរចុងក្រោយក្នុងការផលិត tryptophan គឺ កាតាលីករដោយអង់ស៊ីម tryptophan synthase ដែលត្រូវបានគេស្គាល់ថា tryptophan synthetase ។
Eubacteria, Archaebacteria, Protista, Fungi និង Plantae គឺជាម្ចាស់ផ្ទះញឹកញាប់សម្រាប់វា។ ទោះយ៉ាងណាក៏ដោយ Animalia មិនមានវាទេ។ ជាធម្មតាវាលេចឡើងជា 2 2 tetramer ។
ផ្នែករងជំរុញការបំប្លែងបញ្ច្រាសនៃ indole-3-glycerol phosphate ទៅindole និង glyceraldehyde-3-phosphate (G3P) (IGP) ។
នៅក្នុងដំណើរការដែលពឹងផ្អែកពី pyridoxal phosphate (PLP) អង្គភាពរងជំរុញឱ្យមាន condensation ដែលមិនអាចត្រឡប់វិញនៃ indole និង serine ដើម្បីបង្កើត tryptophan ។
ឆានែល hydrophobic ខាងក្នុងដែលមានប្រវែង 25 angstroms និងមានទីតាំងនៅក្នុងអង់ស៊ីមភ្ជាប់កន្លែងសកម្មនីមួយៗទៅកន្លែងសកម្មដែលនៅជាប់គ្នា។
សូមមើលផងដែរ: តើអ្វីជាភាពខុសគ្នារវាងក្រុមហ៊ុនអន្តរជាតិ និងពហុជាតិ? - ភាពខុសគ្នាទាំងអស់។នេះជំរុញការបញ្ជូនស្រទាប់ខាងក្រោម ដែលជាយន្តការដែល indole ផលិតនៅកន្លែងសកម្ម សាយភាយដោយផ្ទាល់ទៅកាន់គេហទំព័រសកម្មផ្សេងទៀត។ Tryptophan synthase មានទីតាំងសកម្មដែលផ្សំគ្នាយ៉ាងច្រើន។
Eubacteria, Archaebacteria, Protista, Fungi និង Plantae ត្រូវបានគេរកឃើញជាញឹកញាប់ដើម្បីរួមបញ្ចូល tryptophan synthase។ មនុស្សនិងសត្វដទៃទៀតខ្វះវា។
អាស៊ីតអាមីណូមួយក្នុងចំណោមអាស៊ីតអាមីណូចាំបាច់ចំនួនប្រាំបួនសម្រាប់មនុស្ស សារធាតុ tryptophan គឺជាអាស៊ីតអាមីណូស្តង់ដារមួយក្នុងចំណោមអាស៊ីតអាមីណូស្តង់ដារចំនួនម្ភៃ។ ដូច្នេះសារធាតុ Tryptophan គឺចាំបាច់សម្រាប់របបអាហាររបស់មនុស្ស។
វាត្រូវបានគេដឹងផងដែរថា tryptophan synthetase អាចប្រើអាណាឡូក indole ដូចជា fluorinated ឬ methylated indoles ជាស្រទាប់ខាងក្រោមដើម្បីបង្កើតអាណាឡូក tryptophan សមមូល។
Pseudouridine
អក្សរក្រិច psi- ត្រូវបានប្រើដើម្បីអក្សរកាត់ pseudouridine ដែលជា isomer នៃ nucleoside uridine ដែល uracil ត្រូវបានភ្ជាប់ជាមួយអាតូមកាបូនដោយតំណភ្ជាប់កាបូន-កាបូន ជាជាងការតភ្ជាប់ glycosidic អាសូត-កាបូន។ (យូរៗម្តង Uracil ត្រូវបានគេហៅថា "pseudoracil" នៅក្នុងការរៀបចំនេះ។)
RNA រីករាលដាលបំផុតការផ្លាស់ប្តូរនៅក្នុង RNA កោសិកាគឺ pseudouridine ។ RNA អាចឆ្លងកាត់ការបំរែបំរួលគីមីពិសេសជាង 100 កំឡុងពេលចម្លង និងសំយោគ។
បន្ថែមពីលើនុយក្លេអូទីតធម្មតាចំនួនបួន ទាំងនេះអាចប៉ះពាល់ដល់កន្សោម RNA ក្រោយការចម្លង ហើយមានមុខងារមួយចំនួននៅក្នុងកោសិកា រួមទាំងការបកប្រែ RNA ការធ្វើមូលដ្ឋានីយកម្ម និងស្ថេរភាព។
មួយក្នុងចំណោមទាំងនេះគឺ pseudouridine ដែលជាអ៊ីសូម C5-glycoside នៃ uridine ជាមួយនឹងចំណង C-C រវាង C1 នៃជាតិស្ករ ribose និង C5 នៃ uracil ជំនួសឱ្យចំណង C1-N1 ធម្មតាដែលមាននៅក្នុង uridine ។
វាមានភាពចល័តបង្វិលបន្ថែម និងភាពបត់បែនតាមទម្រង់ ដោយសារចំណង C-C ។ លើសពីនេះ ទីតាំង N1 នៃ pseudouridine មានអ្នកផ្តល់ចំណងអ៊ីដ្រូសែនបន្ថែម។
Pseudouridine ហៅផងដែរថា 5-ribosyluracil គឺជាធាតុផ្សំដ៏អាថ៌កំបាំងនៃ RNAs រចនាសម្ព័ន្ធ (ផ្ទេរ ribosomal នុយក្លេអ៊ែរតូច (snRNA)) និង នុយក្លេអ៊ែរតូច) ។ ថ្មីៗនេះវាត្រូវបានគេរកឃើញនៅក្នុងការសរសេរកូដ RNA ផងដែរ។
វាគឺជាការរកឃើញដំបូងគេ គឺជាការរីករាលដាលបំផុត ហើយអាចត្រូវបានរកឃើញនៅក្នុងដែនវិវត្តន៍ទាំងបីនៃជីវិត។ នៅក្នុង tRNA yeast, pseudouridine បង្កើតបានប្រហែល 4% នៃ nucleotides
តាមរយៈការបង្កើតចំណងអ៊ីដ្រូសែនបន្ថែមជាមួយនឹងទឹក ការផ្លាស់ប្តូរមូលដ្ឋាននេះអាចស្ថេរភាព RNA និងបង្កើនការជង់មូលដ្ឋាន។
The ចំនួន pseudouridines កើនឡើងជាមួយនឹងភាពស្មុគស្មាញនៃសារពាង្គកាយ។ មាន 11 pseudouridines នៅក្នុងrRNA នៃ Escherichia coli, 30 នៅក្នុង cytoplasmic rRNA នៃ yeast, ការផ្លាស់ប្តូរមួយនៅក្នុង mitochondrial 21S rRNA និងប្រហែល 100 នៅក្នុង rRNA របស់មនុស្ស។
សូមមើលផងដែរ: Dupont Corian Vs LG Hi-Macs: តើមានភាពខុសគ្នាអ្វីខ្លះ?វាត្រូវបានបង្ហាញថា pseudouridine នៅក្នុង RNA-tunet និង និងធ្វើឱ្យមានស្ថេរភាពនៃរចនាសម្ព័ន្ធក្នុងតំបន់ និងជំនួយក្នុងការថែរក្សាតួនាទីរបស់ពួកគេក្នុងការឌិកូដ mRNA ការជួបប្រជុំគ្នា ribosome ដំណើរការ និងការបកប្រែ។
វាត្រូវបានបង្ហាញថា pseudouridine នៅក្នុង snRNA ធ្វើអោយប្រសើរឡើងនូវចំណុចប្រទាក់រវាង pre-mRNA និង spliceosomal RNA ដើម្បីជួយគ្រប់គ្រងការភ្ជាប់គ្នា។
Fatty Acid Synthase
The FASN ហ្សែននៅក្នុងមនុស្សបានអ៊ិនកូដអង់ស៊ីមដែលគេស្គាល់ថាជាអាស៊ីតខ្លាញ់សំយោគ (FAS) ។ ប្រូតេអ៊ីនពហុអង់ស៊ីមដែលហៅថា fatty acid synthase ជំរុញការសំយោគអាស៊ីតខ្លាញ់។
វាជាប្រព័ន្ធអង់ស៊ីមទាំងមូល មិនមែនត្រឹមតែអង់ស៊ីមមួយទេ ដែលផ្សំឡើងពីសារធាតុ polypeptides ពហុមុខងារ 272 kDa ដូចគ្នាបេះបិទ ដែលផ្ទេរស្រទាប់ខាងក្រោមពីដែនមុខងារមួយទៅដែនមុខងារមួយទៀត។
ការងារចម្បងរបស់វាគឺប្រើប្រាស់ NADPH ដើម្បីជំរុញការបង្កើត palmitate (C16:0 ដែលជាអាស៊ីតខ្លាញ់ឆ្អែតខ្សែសង្វាក់វែង) ពី acetyl- និង malonyl-CoA
Acetyl-CoA និង malonyl -CoA ត្រូវបានបំប្លែងទៅជាអាស៊ីតខ្លាញ់តាមរយៈលំដាប់នៃដំណើរការ decarboxylative Claisen condensation។
បន្ទាប់ពីការពន្លូតជុំនីមួយៗ ketoreductase (KR), dehydratase (DH) និង enoyl reductase ដំណើរការតាមលំដាប់លំដោយដើម្បីបន្ថយក្រុម beta keto ទៅខ្សែសង្វាក់កាបូនដែលឆ្អែតទាំងស្រុង(ER) ។
នៅពេលដែលខ្សែសង្វាក់អាស៊ីតខ្លាញ់បានកើនឡើងដល់ប្រវែង 16 កាបូន វាត្រូវបានបញ្ចេញដោយសកម្មភាពរបស់ thioesterase (TE) ដែលត្រូវបានភ្ជាប់ជាកូឡាទីនទៅនឹងក្រុមសិប្បនិម្មិត phosphopantetheine នៃប្រូតេអ៊ីន acyl carrier protein (ACP) ។ ជាទូទៅគេហៅថា UDP-glucose៖ (1→4) 4-D-glucosyltransferase របស់អង់ស៊ីមសម្រាប់ D-glucan។
GDP-glucose ត្រូវបានប្រើប្រាស់ដោយអង់ស៊ីមដែលទាក់ទងគ្នាហៅថា cellulose synthase (GDP- ការបង្កើត) (EC 2.4.1.29) ។ ទាំងបាក់តេរី និងរុក្ខជាតិមានសមាជិកនៃក្រុមអង់ស៊ីមនេះ។
សមាជិកបាក់តេរីអាចត្រូវបានគេស្គាល់ផងដែរថាជា BcsA (បាក់តេរី cellulose synthase) ឬ CelA ខណៈពេលដែលសមាជិករុក្ខជាតិត្រូវបានគេស្គាល់ជាទូទៅថាជា CesA (cellulose synthase) ឬ CslA (cellulose synthase-like) (សាមញ្ញ "សែលុយឡូស") .
CesA ត្រូវបានទទួលដោយរុក្ខជាតិជាលទ្ធផលនៃ endosymbiosis ដែលបង្កើតដល់ chloroplast ។ គ្រួសារទី 2 នៃ glucosyltransferases រួមមានប្រភេទនេះ (GT2)។
ជីវម៉ាស់ភាគច្រើននៅលើផែនដីត្រូវបានផលិតតាមរយៈការសំយោគ និងអ៊ីដ្រូលីស៊ីសដោយអង់ស៊ីមហៅថា glycosyltransferases។
រុក្ខជាតិ CesA superfamily ត្រូវបានគេស្គាល់ថាមានផ្ទុក អំបូររងចំនួនប្រាំពីរ ហើយក្រុមរុក្ខជាតិ-algal superfamily រួមបញ្ចូលគ្នាមាន 10.
ក្រុមសត្វតែមួយគត់ដែលមានអង់ស៊ីមនេះគឺurochordates ដែលបានទទួលវាតាមរយៈការផ្ទេរហ្សែនផ្តេកជាង 530 លានឆ្នាំមុន។
Cellulose Synthase (GDP-Forming)
អង់ស៊ីមនេះគឺជាសមាជិកនៃក្រុមរង hexosyltransferase នៃ glycosyltransferases ។ ថ្នាក់អង់ស៊ីមនេះត្រូវបានសំដៅដោយឈ្មោះវិទ្យាសាស្ត្ររបស់វា GDP-glucose: 1,4-beta-D-glucan 4-beta-D-glucosyltransferase ។
ឈ្មោះផ្សេងទៀតដែលត្រូវបានគេប្រើញឹកញាប់គឺ cellulose synthase (guanosine diphosphate-forming) cellulose synthetase និង guanosine diphosphoglucose-1,4-beta-glucan glucosyltransferase។ អង់ស៊ីមនេះចូលរួមនៅក្នុងការរំលាយអាហាររបស់ sucrose និងម្សៅ។
អ្វីទៅជា Synthetase?
ពាក្យ "សំយោគ" ដែលជួនកាលគេស្គាល់ថាជា "លីហ្គាស" សំដៅទៅលើប្រភេទណាមួយនៃអង់ស៊ីមប្រហែល 50 ដែលជំរុញឱ្យមានប្រតិកម្មសន្សំថាមពលគីមី និងសម្រុះសម្រួលរវាងព្រឹត្តិការណ៍បែកបាក់ដែលប្រើប្រាស់ថាមពល និងផលិតភាព។ ដំណើរការសំយោគ។
ដោយការបំបែកចំណងផូស្វាតដ៏ស្វាហាប់ ពួកវាបង្កើតថាមពលចាំបាច់ដើម្បីជំរុញការរួមបញ្ចូលគ្នានៃម៉ូលេគុលពីរ (ក្នុងករណីជាច្រើន ដោយការបំប្លែងអាឌីណូស៊ីនទ្រីផូស្វាត [ATP] ទៅជា adenosine diphosphate [ADP]) .
ligase ដែលគេស្គាល់ថាជាអាស៊ីតអាមីណូ-RNA ligase គឺជាសារធាតុមួយដែលជំរុញការបង្កើតចំណងកាបូនអុកស៊ីហ្សែនរវាង RNA ផ្ទេរ និងអាស៊ីតអាមីណូ។
នៅពេលដែលអង់ស៊ីមមួយចំនួនដូចជា amide synthetases និង peptide synthetases សកម្ម កាបូន-nitrogen (C-N)ចំណងត្រូវបានបង្កើតឡើង។
ការសំយោគត្រូវបានគេស្គាល់ផងដែរថាជា Ligase
ភាពខុសគ្នារវាង Synthetase និង Synthase
សំយោគតេតាសគឺជាអង់ស៊ីមដែលអាចជំរុញការភ្ជាប់នៃ ម៉ូលេគុលធំពីរដោយបង្កើតចំណងគីមីថ្មីមួយ ជាធម្មតាជាមួយនឹងអ៊ីដ្រូលីស៊ីសក្នុងពេលដំណាលគ្នានៃក្រុមគីមី pendant តូចមួយនៅលើម៉ូលេគុលធំមួយ ឬវាអាចជំរុញការភ្ជាប់នៃសមាសធាតុពីរដូចជាការភ្ជាប់ C-O, C-S, C-N ជាដើម។
A ligase ជាធម្មតាបណ្តាលឱ្យមានប្រតិកម្មដូចខាងក្រោមកើតឡើង៖
- A-C + b = Ab + C
- A+D + B + C + D + E + F = Ab + cD
នៅកន្លែងដែលអាស្រ័យ ការដាក់ក្រុមតូចៗត្រូវបានតំណាងដោយអក្សរតូច។ Ligase អាចជួសជុលការបំបែកខ្សែតែមួយដែលវិវឌ្ឍន៍នៅក្នុង DNA ខ្សែពីរកំឡុងពេលចម្លង ក៏ដូចជាភ្ជាប់បំណែកអាស៊ីត nucleic ពីរដែលបំពេញបន្ថែម។
ម្យ៉ាងវិញទៀត synthase គឺជាអង់ស៊ីមដែលជំរុញដំណើរការសំយោគនៅក្នុងជីវគីមី។ ពួកគេត្រូវបានរួមបញ្ចូលនៅក្នុងប្រភេទនៃ lyases យោងតាមការចាត់ថ្នាក់លេខ EC ។
Nomenclature
សូមចាំថា នាមនាមជីវសាស្រ្តដំបូងត្រូវបានបែងចែករវាងសំយោគ និងសំយោគ។ អនុលោមតាមនិយមន័យដើម ការសំយោគប្រើប្រាស់ nucleoside triphosphates (ដូចជា ATP, GTP, CTP, TTP និង UTP) ជាប្រភពថាមពលខណៈពេលដែលសំយោគមិនមាន។
ទោះជាយ៉ាងនេះក្តី យោងទៅតាមគណៈកម្មការចម្រុះស្តីពីឈ្មោះគីមីជីវៈ (JCBN) "សំយោគ"