Каква е разликата между синтаза и синтеза? (Разкрити факти) - Всички разлики

Съдържание
Класът лигазни ензими E.C. 6 включва синтетази и синтетази. Те участват в синтетични реакции и катализират комбинирането на две молекули, като едновременно с това разкъсват дифосфатната връзка в АТФ или друг сравним трифосфат.
За разлика от синтетазите, които използват АТФ като източник на енергия по време на синтеза на биологични съединения, синтетазите са всякакви лигази, които катализират синтеза на биологични съединения, без да използват АТФ като източник на енергия.
В тази статия ще научите каква е разликата между синтеза и синтеза.
Какво представлява синтазата?
Синтазата е ензим, който катализира процеса на синтез в биохимията. Имайте предвид, че в биологичната номенклатура първоначално се прави разлика между синтези и синтези.
В съответствие с първоначалното определение синтетазите използват нуклеозидните трифосфати (като АТФ, ГТФ, ЦТФ, ТТФ и УТФ) като източник на енергия, докато синтетазите не го правят.
Въпреки това според Съвместната комисия по биохимична номенклатура (JCBN) "синтаза" може да се използва за обозначаване на всеки ензим, който катализира синтеза (независимо дали използва нуклеозидтрифосфати), но "синтетаза" трябва да се използва само за обозначаване на "лигаза".
Ето списък с примери за различни видове синтази:
- АТФ синтаза
- Цитрат синтаза
- Триптофан синтаза
- Псевдоуридинова синтаза
- Синтаза на мастните киселини
- Синтаза на целулозата (UDP-образуваща)
- Синтаза на целулозата (образуваща БВП)
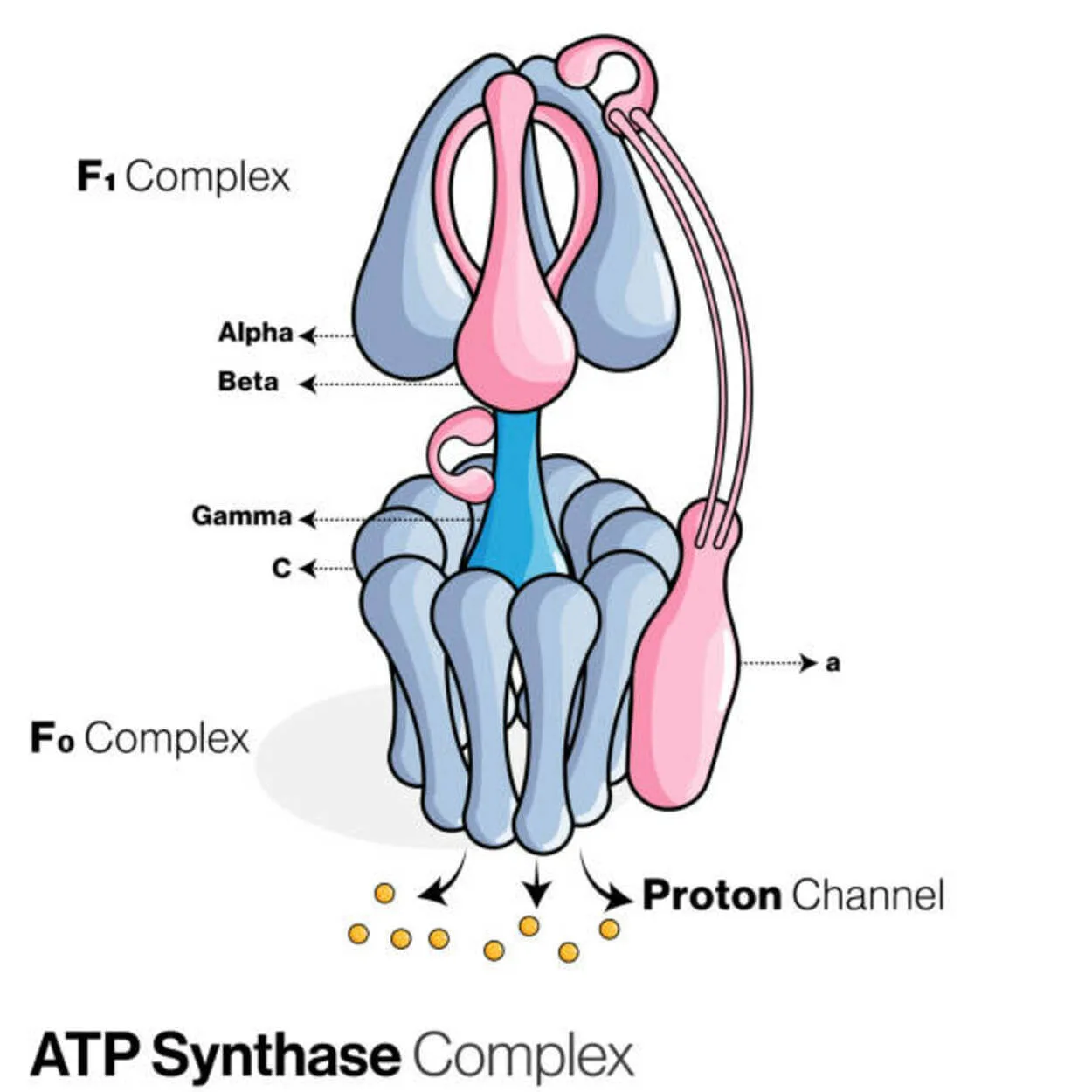
АТФ синтаза
Аденозин дифосфатът (АДФ) и неорганичният фосфат се използват за създаване на молекулата за съхранение на енергия аденозин трифосфат (АТФ) от протеин, наречен АТФ синтаза (Pi).
Категоризиран е като лигаза, тъй като модифицира ADP чрез образуване на P-O връзка (фосфодиестерна връзка). Молекулярно устройство, наречено АТФ-синтаза.
От енергийна гледна точка производството на АТФ от ADP и Pi е нежелателно и процесът обикновено протича в обратна посока.
Концентрационният градиент на протоните (H+) през вътрешната митохондриална мембрана при еукариотите или плазмената мембрана при бактериите задвижва тази реакция, като свързва синтеза на АТФ по време на клетъчното дишане с градиента.
В растенията АТФ-синтазата използва протонния градиент, образуван в тилакоидния лумен, през тилакоидната мембрана и в стромата на хлоропласта, за да произвежда АТФ по време на фотосинтезата.
За АТФ-аза еукариотните АТФ-синтетази са F-ATPases, които функционират "в обратна посока." Този вид е основно разгледан в тази статия. FO и F1 субединиците на F-ATPase имат ротационен моторен механизъм, който позволява синтеза на АТФ.
Съществуват различни видове синтаза
Цитрат синтаза
Почти всички живи клетки съдържат ензима цитрат синтетаза, който служи като пейсмейкър в първата стъпка на цикъла на лимонената киселина и е обозначен като E.C. 2.3.3.1 (преди 4.1.3.7). (или цикъл на Кребс).
Цитрат синтетазата се намира в митохондриалния матрикс на еукариотните клетки, въпреки че ядрената ДНК, а не митохондриалната ДНК, я кодира.
Вижте също: Индийци срещу пакистанци (основни разлики) - Всички разликиТой се създава в цитоплазмата от цитоплазмените рибозоми и впоследствие се премества в митохондриалния матрикс.
Типичен количествен ензимен маркер за наличието на интактни митохондрии е цитрат синтазата. Пиковата активност на цитрат синтазата показва колко митохондрии има в скелетните мускули.
Високоинтензивното интервално обучение има потенциала да повиши максималната активност повече, отколкото обучението за издръжливост или високоинтензивното интервално обучение.
Ацетилкоензим А има двувъглероден ацетатен остатък и молекула от четиривъглероден оксалоацетат се кондензира, за да се получи шествъглероден цитрат, който се получава чрез реакцията на кондензация, катализирана от цитратсинтазата.
Триптофан синтаза
Последните два етапа от производството на триптофан се катализират от ензима триптофан синтаза, известен също като триптофан синтетаза.
Вижте също: Каква е разликата между млякото с витамин D и пълномасленото мляко? (Обяснено) - Всички разликиЧести негови гостоприемници са Eubacteria, Archaebacteria, Protista, Fungi и Plantae. В Animalia обаче той не се среща. Обикновено се появява като 2 2 тетрамер.
Субединиците катализират обратимото превръщане на индол-3-глицерол фосфат в индол и глицералдехид-3-фосфат (G3P) (IGP).
В процеса, зависим от пиридоксалфосфат (PLP), субединиците катализират необратимата кондензация на индол и серин, за да се получи триптофан.
Вътрешен хидрофобен канал с дължина 25 ангстрьома, разположен в ензима, свързва всеки активен сайт със съседния активен сайт.
Това спомага за канализирането на субстрата - механизъм, при който индолът, произведен в активните места, дифундира директно към други активни места. Триптофан синтазата съдържа алостерично свързани активни места.
В Eubacteria, Archaebacteria, Protista, Fungi и Plantae често се открива триптофан синтаза. Хората и другите животни я нямат.
Една от деветте необходими за човека аминокиселини, триптофанът е една от двадесетте стандартни аминокиселини. Следователно триптофанът е от съществено значение за човешката диета.
Известно е също така, че триптофансинтетазата може да използва индолови аналози, като флуорирани или метилирани индоли, като субстрати за получаване на еквивалентни триптофанови аналози.
Псевдоуридин
Гръцката буква psi- се използва за съкращаване на псевдоуридин, изомер на нуклеозида уридин, при който урацилът е свързан с въглеродния атом чрез въглеродно-въглеродна връзка, а не чрез азотно-въглеродна гликозидна връзка (понякога урацилът се нарича "псевдоурацил" в този вид).
Най-разпространеното изменение на РНК в клетъчната РНК е псевдоуридинът. РНК може да претърпи над 100 уникални в химично отношение изменения по време на транскрипцията и синтеза.
В допълнение към четирите конвенционални нуклеотида, те могат потенциално да повлияят на експресията на РНК посттранскрипционно и да имат редица функции в клетката, включително транслация, локализация и стабилност на РНК.
Един от тях е псевдоуридинът - С5-гликозиден изомер на уридина с връзка С-С между С1 на рибозната захар и С5 на урацила вместо типичната за уридина връзка С1-Н1.
Той има допълнителна ротационна подвижност и конформационна гъвкавост поради връзката С-С. Освен това позицията N1 на псевдоуридина притежава допълнителен донор на водородна връзка.
Псевдоуридинът, наричан още 5-рибозилурацил, е познат, но загадъчен компонент на структурните РНК (трансферна, рибозомна, малка ядрена (snRNA) и малка ядрена). Наскоро той беше открит и в кодиращата РНК.
Той е първият, който е открит, най-разпространен е и може да бъде открит във всички три еволюционни области на живота. В тРНК на дрождите псевдоуридинът съставлява около 4 % от нуклеотидите.
Чрез образуването на допълнителни водородни връзки с водата тази промяна на базата може да стабилизира РНК и да подобри подреждането на базите.
Броят на псевдоуридините нараства със сложността на организма. 11 псевдоуридини има в рРНК на Escherichia coli, 30 в цитоплазмената рРНК на дрожди, една промяна в митохондриалната 21S рРНК и около 100 в рРНК на човека.
Доказано е, че псевдоуридинът в рРНК и тРНК прецизира и стабилизира регионалната структура и спомага за поддържането на ролята им в декодирането на мРНК, сглобяването на рибозомите, обработката и транслацията.
Доказано е, че псевдоуридинът в snRNA подобрява връзката между пре-мРНК и сплайсозомната РНК, за да подпомогне регулирането на сплайсинга.
Синтаза на мастните киселини
Генът FASN при хората кодира ензима, известен като синтеза на мастни киселини (FAS). Мултиензимен протеин, наречен синтез на мастни киселини, катализира синтеза на мастни киселини.
Това е цяла ензимна система, а не само един ензим, състоящ се от два идентични многофункционални полипептида с размер 272 kDa, които прехвърлят субстрати от един функционален домейн към друг.
Основната му задача е да използва NADPH, за да катализира създаването на палмитат (C16:0, наситена мастна киселина с дълга верига) от ацетил- и малонил-КоА
Ацетил-КоА и малонил-КоА се превръщат в мастни киселини чрез поредица от декарбоксилационни процеси на кондензация по Клайзен.
След всеки цикъл на удължаване кеторедуктаза (KR), дехидратаза (DH) и еноилредуктаза работят последователно, за да намалят бета кетогрупата до напълно наситена въглеродна верига (ER).
Когато веригата на мастната киселина достигне дължина от 16 въглеродни атома, тя се освобождава под действието на тиоестераза (ТЕ), която е ковалентно свързана с фосфопантеетиновата протетична група на ацил-носителя (АСР) (палмитинова киселина).
Синтаза на целулозата (UDP-формиране)
Основният ензим, отговорен за производството на целулоза, е целулозната синтетаза (EC 2.4.1.12) в нейната UDP-формираща форма. Обикновено тя се нарича UDP-глюкоза: (1→4) Ензимология 4-D-глюкозилтрансфераза за D-глюкан.
GDP-глюкозата се използва от сроден ензим, наречен целулозна синтетаза (образуваща GDP) (EC 2.4.1.29). И бактериите, и растенията имат членове на това семейство ензими.
Бактериалните членове могат да бъдат известни и като BcsA (бактериална целулозна синтаза) или CelA, докато растителните членове обикновено са известни като CesA (целулозна синтаза) или спекулативното CslA (подобна на целулозна синтаза) (просто "целулоза").
CesA е придобита от растенията в резултат на ендосимбиозата, довела до появата на хлоропласта. Семейство 2 на глюкозилтрансферазите включва тази (GT2).
По-голямата част от биомасата на Земята се произвежда чрез биосинтеза и хидролиза от ензими, наречени гликозилтрансферази.
Известно е, че растителното суперсемейство на CesA съдържа седем подсемейства, а комбинираното суперсемейство на растенията и водораслите - 10.
Единствената животинска група, която притежава този ензим, са урохордите, които са го получили чрез хоризонтален трансфер на гени преди повече от 530 милиона години.
Синтаза на целулозата (формираща БВП)
Този ензим е член на подсемейството на хексозилтрансферазите от групата на гликозилтрансферазите. Този клас ензими се нарича с научното си наименование GDP-глюкоза:1,4-бета-D-глюкан 4-бета-D-глюкозилтрансфераза.
Други имена, които често се използват, са целулозна синтетаза (гуанозин дифосфат-формираща), целулозна синтетаза и гуанозин дифосфоглюкозо-1,4-бета-глюкан глюкозилтрансфераза. Този ензим взема участие в метаболизма на захарозата и нишестето.
Какво представлява синтетазата?
Терминът "синтетаза", понякога известен като "лигаза", се отнася за всеки един от клас от около 50 ензима, които катализират химични реакции, запазващи енергията, и посредничат между енергоемки разпадни процеси и продуктивни синтетични процеси.
Като разцепват енергийна фосфатна връзка, те генерират необходимата енергия за катализиране на комбинирането на две молекули (в много случаи чрез едновременно превръщане на аденозинтрифосфат [ATP] в аденозиндифосфат [ADP]).
Лигаза, известна като лигаза аминокиселини-РНК, е тази, която катализира създаването на въглеродно-кислородна връзка между трансферна РНК и аминокиселина.
Когато някои ензими, като например амидните синтетази и пептидните синтетази, са активни, се образуват въглеродно-азотни (C-N) връзки.
Синтетазата е известна и като лигаза
Разлика между синтаза и синтеза
Синтетазата е ензим, който може да катализира свързването на две големи молекули чрез създаване на нова химична връзка, обикновено с едновременна хидролиза на малка висяща химична група на една от по-големите молекули, или може да катализира свързването на две съединения, като например свързването на C-O, C-S, C-N и т.н.
Една лигаза обикновено предизвиква следната реакция:
- A-C + b = Ab + C
- A+D + B + C + D + E + F = Ab + cD
В случаите, когато зависимите, малки групировки са представени с малки букви. Лигазата може да поправя едноверижни скъсвания, които се получават в двойноверижната ДНК по време на репликация, както и да свързва два комплементарни фрагмента нуклеинова киселина.
От друга страна, синтазата е ензим, който катализира процеса на синтез в биохимията. Те са включени в категорията на лиазите според категоризацията на EC номера.
Номенклатура
Имайте предвид, че в биологичната номенклатура първоначално се прави разграничение между синтетази и синтетази. В съответствие с първоначалното определение синтетазите използват нуклеозидните трифосфати (като АТФ, ГТФ, ЦТФ, ТТФ и УТФ) като източник на енергия, докато синтетазите не го правят.
Въпреки това според Съвместната комисия по биохимична номенклатура (JCBN) "синтаза" може да се използва за обозначаване на всеки ензим, който катализира синтеза (независимо дали използва нуклеозидтрифосфати), но "синтетаза" трябва да се използва само за обозначаване на "лигаза".
Основното разграничение между Синтаза и Синтетаза е, че Синтетаза е семейство ензими, които могат да създават връзки между молекулите, докато Синтаза е ензим.
| Синтаза | Синтетаза |
| Катализира синтетичен процес без АТФ | Изисква ATP |
| Попада в класификацията за ваза или трансфер | Попада в класификацията на лигазите |
| напр. HMG-COA синтаза, АТФ синтаза | напр. сукцини1-СОА синтетаза, глутамин синтетаза |
Таблица за сравнение на Synthase vs Synthetase
Гледайте този видеоклип, за да научите повече за синтазата срещу синтезата
Заключение
- Синтетазите не се нуждаят от NTP, за да функционират, тъй като те катализират синтетични процеси, които изискват хидролиза на нуклеозидни трифосфати (а не просто ATP).
- През 80-те години на миналия век Комитетът по номенклатура на Международния съюз по биохимия променя определението за синтеза, за да обхване всички синтетични ензими, независимо дали използват НТП, или не, и синтетазата става синоним на лигаза.
- Лигазата е ензим, който съединява две по-малки молекули, като използва енергията от хидролизата на NTP (обикновено чрез реакция на кондензация).