Hvad er forskellen mellem en syntase og en syntetase (fakta afsløret) - Alle forskellene

Indholdsfortegnelse
E.C. 6-klassen af ligaseenzymer omfatter synthaser og synthetaser. De deltager i syntetiske reaktioner og katalyserer kombinationen af to molekyler, samtidig med at de bryder diphosphatforbindelsen i ATP eller et andet tilsvarende trifosfat.
I modsætning til synthetaser, der anvender ATP som energikilde under syntesen af biologiske forbindelser, er synthaser alle ligaser, der katalyserer syntesen af biologiske forbindelser uden at anvende ATP som energikilde.
I denne artikel vil du lære præcis forskellen mellem Synthase og Synthetase at kende.
Hvad er synthase?
En synthase er et enzym, der katalyserer synteseprocessen i biokemien. Husk, at den biologiske nomenklatur oprindeligt skelnede mellem synthetaser og synthaser.
I overensstemmelse med den oprindelige definition anvender synthetaser nukleosidtriphosphater (såsom ATP, GTP, CTP, TTP og UTP) som energikilde, mens synthaser ikke gør det.
Ikke desto mindre kan "synthase" ifølge Joint Commission on Biochemical Nomenclature (JCBN) anvendes til at henvise til ethvert enzym, der katalyserer syntese (uanset om det anvender nukleosidtriphosphater eller ej), men "synthetase" bør kun anvendes til at henvise til "ligase".
Her er en liste med eksempler på forskellige typer af synthaser:
- ATP-syntase
- Citrat-syntase
- Tryptofansyntase
- Pseudouridinsyntase
- Fedtsyresyntase
- Cellulosesyntase (UDP-dannende)
- Cellulosesyntase (GDP-dannende)
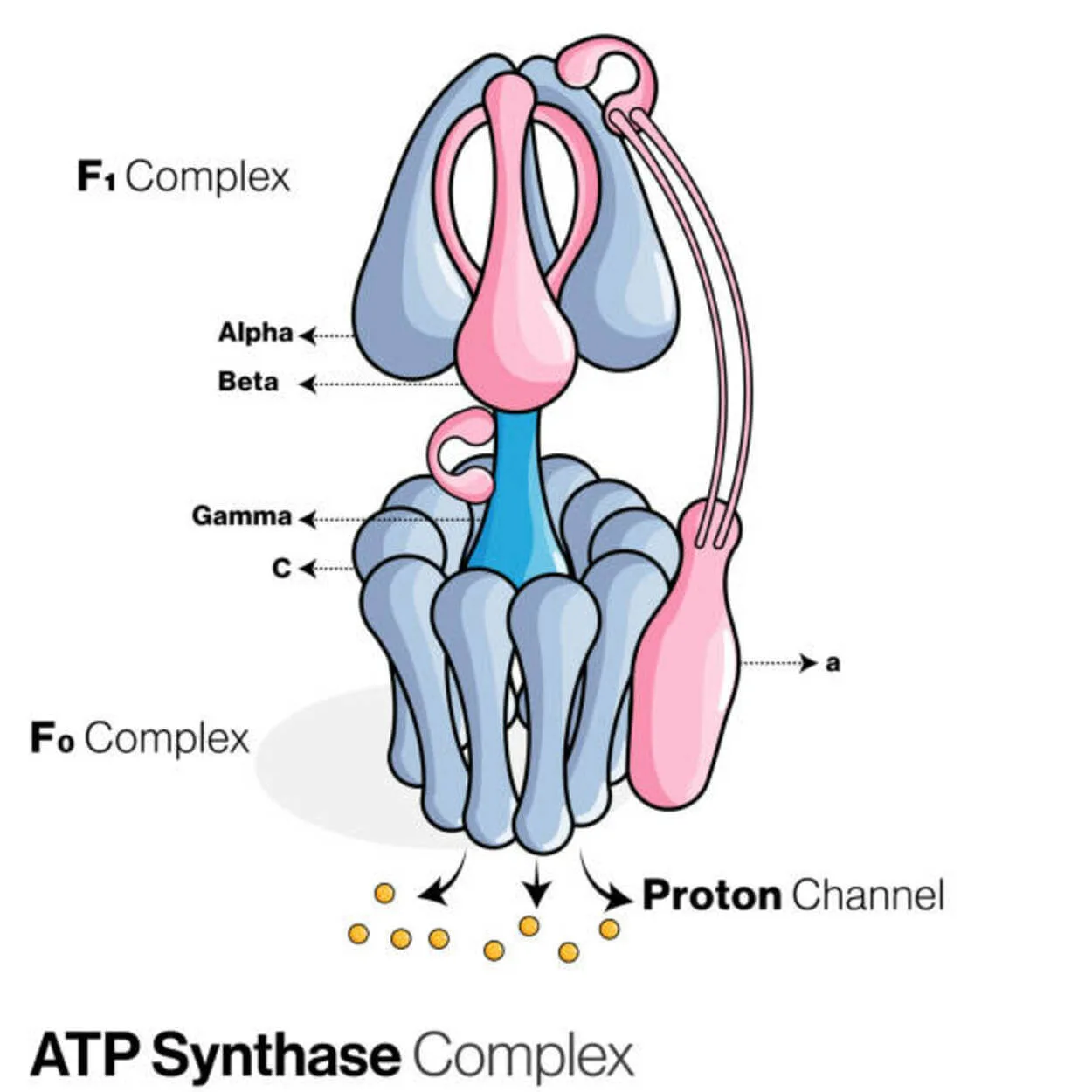
ATP-syntase
Adenosindiphosphat (ADP) og uorganisk fosfat bruges til at skabe energilagringsmolekylet adenosintrifosfat (ATP) af et protein kaldet ATP-syntase (Pi).
Den er kategoriseret som en ligase, da den modificerer ADP ved at danne et P-O-link (phosphodiesterbinding). En molekylær enhed kaldet ATP-syntase.
Energimæssigt er produktionen af ATP fra ADP og Pi uønsket, og processen ville typisk gå den anden vej.
En koncentrationsgradient af protoner (H+) på tværs af den indre mitokondriemembran i eukaryoter eller plasmamembranen i bakterier driver denne reaktion fremad ved at koble ATP-syntesen under celleatmningen til gradienten.
I planter bruger ATP-syntase en protongradient, der dannes i thylakoidlumen, til at producere ATP under fotosyntesen ved hjælp af en protongradient, der dannes i thylakoidlumen over thylakoidmembranen og ind i chloroplaststromaet.
For en ATPase er eukaryote ATP-syntaser F-ATPaser, der fungerer "omvendt". Denne type er primært behandlet i denne artikel. FO- og F1-underenhederne i en F-ATPase har en roterende motormekanisme, der muliggør ATP-syntese.
Der findes forskellige typer af synthase
Citrat-syntase
Næsten alle levende celler indeholder enzymet citratsyntase, der fungerer som pacemaker i første trin af citronsyrecyklussen og betegnes E.C. 2.3.3.3.1 (tidligere 4.1.3.7). (eller Krebs-cyklus).
Se også: Hvad er forskellen mellem Which og Wich (deres betydning) - Alle forskelleCitratsyntase er placeret i den mitokondrielle matrix i eukaryote celler, selv om det er kerne-DNA, ikke mitokondrie-DNA, der koder for den.
Det oprettes i cytoplasmaet af cytoplasmatiske ribosomer og flyttes derefter ind i mitokondriernes matrix.
En typisk kvantitativ enzymmarkør for tilstedeværelsen af intakte mitokondrier er citratsyntase. Citratsyntaseaktivitetens topaktivitet afslører, hvor mange mitokondrier der er til stede i skeletmuskulaturen.
Intervaltræning med høj intensitet har potentiale til at øge den maksimale aktivitet mere end udholdenhedstræning eller intervaltræning med høj intensitet.
Acetylcoenzym A har to kulstofacetatrester og et molekyle af fire kulstofoxaloacetat kondenserer for at generere seks kulstofcitrat, som produceres ved kondensationsreaktionen, der katalyseres af citratsyntase.
Tryptofansyntase
De sidste to trin i produktionen af tryptofan katalyseres af enzymet tryptofansyntase, også kendt som tryptofansyntase.
Eubacteria, Archaebacteria, Protista, Fungi og Plantae er hyppige værter for den. Animalia har den dog ikke. Normalt optræder den som en 2 2 2-tetramer.
Underenhederne katalyserer den reversible omdannelse af indol-3-glycerolphosphat til indol og glyceraldehyd-3-fosfat (G3P) (IGP).
I en pyridoxalphosphat (PLP)-afhængig proces katalyserer underenhederne den irreversible kondensation af indol og serin for at generere tryptofan.
En intern hydrofobisk kanal, der er 25 angstromer lang og placeret i enzymet, forbinder hvert aktivt sted med det tilstødende aktive sted.
Dette fremmer substratkanalisering, en mekanisme, hvorved indol produceret på aktive steder diffunderer direkte til andre aktive steder. Tryptophansyntase indeholder allosterisk koblede aktive steder.
Eubakterier, Archaebacteria, Protista, Fungi og Plantae indeholder ofte tryptofansyntase, mens mennesker og andre dyr mangler den.
Tryptofan er en af de ni nødvendige aminosyrer for mennesker og er en af de tyve standardaminosyrer. Tryptofan er derfor uundværlig i den menneskelige kost.
Det er også kendt, at tryptophansyntase kan bruge indol-analoger, såsom fluorerede eller methylerede indoler, som substrater til at producere de tilsvarende tryptophananaloger.
Pseudouridin
Det græske bogstav psi- bruges til at forkortes pseudouridin, en isomer af nukleosidet uridin, hvor uracil er forbundet til kulstofatomet ved hjælp af en kulstof-kulstofforbindelse i stedet for en nitrogen-kulstof-glykosidisk forbindelse (Uracil omtales undertiden som "pseudouracil" i denne opstilling).
Den mest udbredte RNA-ændring i cellulært RNA er pseudouridin. RNA kan undergå over 100 kemisk unikke ændringer under transkription og syntese.
Ud over de fire konventionelle nukleotider kan disse potentielt påvirke RNA-ekspressionen posttransskriptionelt og have en række funktioner i cellen, herunder RNA-oversættelse, lokalisering og stabilitet.
En af disse er pseudouridin, en C5-glykosidisomer af uridin med en C-C-binding mellem ribosesukkerens C1 og uracils C5 i stedet for den typiske C1-N1-binding, der findes i uridin.
Det har yderligere rotationsmobilitet og konformationsfleksibilitet på grund af C-C-bindingen. Desuden har pseudouridinets N1-position en ekstra hydrogenbindingsdonor.
Pseudouridin, også kaldet 5-ribosyluracil, er en velkendt, men mystisk komponent i strukturelt RNA (transfer-, ribosomalt, lille nukleært (snRNA) og lille nukleolært RNA). Det blev for nylig også fundet i kodende RNA.
Det var det første, der blev opdaget, er det mest udbredte og findes i alle tre evolutionære områder af livet. I gærens tRNA udgør pseudouridin ca. 4 % af nukleotiderne.
Gennem dannelsen af yderligere hydrogenbindinger med vand er denne baseændring i stand til at stabilisere RNA og forbedre base-stacking.
Antallet af pseudouridiner vokser med organismens kompleksitet. Der er 11 pseudouridiner i Escherichia coli's rRNA, 30 i gærens cytoplasmatiske rRNA, én ændring i mitokondriens 21S rRNA og omkring 100 i menneskets rRNA.
Det er blevet påvist, at pseudouridin i rRNA og tRNA finjusterer og stabiliserer den regionale struktur og bidrager til opretholdelsen af deres roller i mRNA-afkodning, ribosom samling, behandling og oversættelse.
Det er blevet påvist, at pseudouridin i snRNA forbedrer grænsefladen mellem pre-mRNA og det splejsosomale RNA for at hjælpe med at regulere splejsning.
Fedtsyresyntase
FASN-genet hos mennesker koder for et enzym kaldet fedtsyresyntase (FAS). Et multienzymprotein kaldet fedtsyresyntase katalyserer syntesen af fedtsyrer.
Det er et helt enzymatisk system, ikke bare et enkelt enzym, der består af to identiske 272 kDa multifunktionelle polypeptider, som overfører substrater fra et funktionelt domæne til det næste.
Dens primære opgave er at bruge NADPH til at katalysere dannelsen af palmitat (C16:0, en langkædet mættet fedtsyre) fra acetyl- og malonyl-CoA
Acetyl-CoA og malonyl-CoA omdannes til fedtsyrer gennem en række decarboxylative Claisen-kondensationsprocesser.
Efter hver forlængelsesrunde arbejder en ketoreduktase (KR), en dehydratase (DH) og enoylreduktase i rækkefølge for at reducere betaketogruppen til den fuldstændigt mættede kulstofkæde (ER).
Når fedtsyrekæden er vokset til en længde på 16 kulstofatomer, frigøres den ved hjælp af en thioesterase (TE), som er kovalent forbundet med fosfopantetheinprotetikumgruppen i et acylbærerprotein (ACP) (palmitinsyre).
Cellulosesyntase (UDP-dannende)
Det primære enzym, der er ansvarlig for produktionen af cellulose, er cellulosesyntase (EC 2.4.1.12) i sin UDP-formende form. Det kaldes generelt UDP-glucose: (1→4) Enzymologiens 4-D-glucosyltransferase for D-glucan.
GDP-glukose anvendes af et beslægtet enzym kaldet cellulosesyntase (GDP-dannende) (EC 2.4.1.29). Både bakterier og planter har medlemmer af denne enzymfamilie.
Bakterielle medlemmer kan også være kendt som BcsA (bakteriel cellulosesyntase) eller CelA, mens plantemedlemmer typisk er kendt som CesA (cellulosesyntase) eller det spekulative CslA (cellulosesyntase-lignende) (blot "cellulose").
CesA blev erhvervet af planter som følge af den endosymbiose, der gav anledning til kloroplasten. Glucosyltransferaser i familie 2 omfatter denne glucosyltransferase (GT2).
Størstedelen af biomassen på Jorden produceres gennem biosyntese og hydrolyse af enzymer kaldet glycosyltransferaser.
Plante CesA-superfamilien er kendt for at indeholde syv underfamilier, og den kombinerede plante-alge-superfamilie indeholder 10 underfamilier.
Den eneste dyregruppe, der har dette enzym, er urokordaterne, som fik det gennem horisontal genoverførsel for mere end 530 millioner år siden.
Cellulosesyntase (GDP-dannende)
Dette enzym er medlem af hexosyltransferase-underfamilien af glykosyltransferaser. Denne enzymklasse kaldes ved sit videnskabelige navn GDP-glucose:1,4-beta-D-glucan 4-beta-D-glukosyltransferase.
Andre navne, der ofte anvendes, er cellulosesyntase (guanosindiphosphatdannende), cellulosesyntetase og guanosindiphosphoglucose-1,4-beta-glucan-glucosyltransferase. Dette enzym er involveret i metabolismen af saccharose og stivelse.
Hvad er synthetase?
Udtrykket "synthetase", undertiden kendt som "ligase", henviser til en klasse af ca. 50 enzymer, der katalyserer kemiske energibesparende reaktioner og formidler mellem energiforbrugende nedbrydningsreaktioner og produktive syntetiske processer.
Ved at spalte en energirigtig fosfatbinding genererer de den nødvendige energi til at katalysere kombinationen af to molekyler (i mange tilfælde ved den samtidige omdannelse af adenosintrifosfat [ATP] til adenosindiphosphat [ADP]).
En ligase, der er kendt som en aminosyre-RNA-ligase, er en ligase, der katalyserer dannelsen af en kulstof-syre-binding mellem et transfer-RNA og en aminosyre.
Se også: Hvad er forskellen mellem en Tesla Super Charger og en Tesla Destination Charger (omkostninger & forskelle forklaret) - Alle forskelleNår visse enzymer, såsom amidsyntetaser og peptidsyntaser, er aktive, dannes der kulstof-stikstof-bindinger (C-N-bindinger).
Synthetase er også kendt som Ligase
Forskellen mellem syntetase og syntase
En synthetase er et enzym, der kan katalysere sammenføjningen af to store molekyler ved at skabe en ny kemisk binding, typisk med samtidig hydrolyse af en lille kemisk sidegruppe på et af de større molekyler, eller det kan katalysere sammenføjningen af to forbindelser, f.eks. sammenføjning af C-O, C-S, C-N osv.
En ligase forårsager typisk følgende reaktion:
- A-C + b = Ab + C
- A+D + B + C + D + E + F = Ab + cD
Hvor de afhængige, små grupperinger er repræsenteret ved små bogstaver. Ligase kan reparere enkeltstrengsbrud, der opstår i dobbeltstrenget DNA under replikationen, samt sammenkæde to komplementære nukleinsyrefragmenter.
På den anden side er synthase et enzym, der katalyserer synteseprocessen i biokemi. De er inkluderet i kategorien lyaser i henhold til EC-nummerkategoriseringen.
Nomenklatur
Husk på, at den biologiske nomenklatur oprindeligt skelnede mellem synthetaser og synthaser. I overensstemmelse med den oprindelige definition anvender synthetaser nukleosidtriphosphater (såsom ATP, GTP, CTP, TTP og UTP) som energikilde, mens synthaser ikke gør det.
Ikke desto mindre kan "synthase" ifølge Joint Commission on Biochemical Nomenclature (JCBN) anvendes til at henvise til ethvert enzym, der katalyserer syntese (uanset om det anvender nukleosidtriphosphater eller ej), men "synthetase" bør kun anvendes til at henvise til "ligase".
Den primære forskel mellem Synthase og Synthetase er, at Synthetase er en familie af enzymer, der kan skabe bindinger mellem molekyler, mens Synthase er et enzym.
| Synthase | Synthetase |
| Katalyserer en syntetisk proces uden ATP | Kræver ATP |
| Hører under klassificeringen vase eller transferase | hører under klassificeringen af ligase |
| f.eks. HMG-COA-syntase, ATP-syntase | f.eks. Succiny1-COA-syntetase, glutamin synthetase |
Sammenligningstabel Synthase vs Synthetase
Se denne video for at få mere at vide om synthase vs. synthetase
Konklusion
- Synthetaser har ikke brug for NTP'er for at fungere, da de katalyserede syntetiske processer, der krævede hydrolyse af nukleosidtriphosphater (ikke blot ATP).
- Den internationale union for biokemis nomenklaturkomité ændrede definitionen af synthase i 1980'erne til at omfatte alle syntetiske enzymer, uanset om de anvender NTP'er eller ej, og synthetase blev synonymt med ligase.
- Ligase er et enzym, der sammenføjer to mindre molekyler ved at udnytte energien fra NTP-hydrolyse (normalt gennem en kondensationsreaktion).