อะไรคือความแตกต่างระหว่าง Synthase และ Synthetase? (เปิดเผยข้อเท็จจริง) – ความแตกต่างทั้งหมด

สารบัญ
เอนไซม์ลิเกสระดับ E.C. 6 ประกอบด้วยซินเทสและซินทีเทส พวกมันมีส่วนร่วมในปฏิกิริยาสังเคราะห์และกระตุ้นการรวมตัวของสองโมเลกุลในขณะเดียวกันก็ทำลายการเชื่อมโยงไดฟอสเฟตใน ATP หรือไตรฟอสเฟตอื่นที่เทียบเคียงได้
ตรงกันข้ามกับซินทีเทสซึ่งใช้ ATP เป็นแหล่งพลังงานในระหว่างการสังเคราะห์ ของสารประกอบทางชีวภาพ การสังเคราะห์คือเอ็นไซม์ใดๆ ที่กระตุ้นการสังเคราะห์สารประกอบทางชีวภาพโดยไม่ใช้ ATP เป็นแหล่งพลังงาน
ในบทความนี้ คุณจะได้ทราบความแตกต่างอย่างชัดเจนระหว่างซินเทสและซินเทเทส
ซินเทสคืออะไร?
ซินเทสเป็นเอนไซม์ที่กระตุ้นกระบวนการสังเคราะห์ทางชีวเคมี โปรดจำไว้ว่าศัพท์เฉพาะทางชีววิทยานั้นแยกความแตกต่างระหว่าง synthetases และ synthases
ตามคำจำกัดความเดิม สารสังเคราะห์ใช้นิวคลีโอไซด์ไตรฟอสเฟต (เช่น ATP, GTP, CTP, TTP และ UTP) เป็นแหล่งพลังงานในขณะที่สังเคราะห์ไม่ได้
อย่างไรก็ตาม ตามรายงานของ Joint Commission on Biochemical Nomenclature (JCBN) "ซินเทส" สามารถใช้เพื่ออ้างถึงเอ็นไซม์ใดๆ ที่กระตุ้นการสังเคราะห์ (ไม่ว่าจะใช้นิวคลีโอไซด์ไตรฟอสเฟตหรือไม่ก็ตาม) แต่ "ซินเทเทส ” ควรใช้เพื่ออ้างถึง “ligase” เท่านั้น
นี่คือรายการตัวอย่างของซินเทสประเภทต่างๆ:
- ATP synthase
- ซิเตรตซินเทส
- โพรไบโอสามารถใช้เพื่ออ้างถึงเอนไซม์ใดๆ ที่กระตุ้นการสังเคราะห์ (ไม่ว่าจะใช้ nucleoside triphosphates หรือไม่) แต่ควรใช้ "synthetase" เพื่ออ้างถึง "ligase" เท่านั้น
ความแตกต่างหลักระหว่าง Synthase และ Synthetase คือ ว่า Synthetase เป็นตระกูลของเอนไซม์ที่อาจสร้างพันธะระหว่างโมเลกุล ในขณะที่ Synthase เป็นเอนไซม์
Synthase Synthetase กระตุ้นกระบวนการสังเคราะห์โดยปราศจาก ATP ต้องการ ATP จัดอยู่ในการจัดประเภทแจกันหรือทรานสเฟอร์เรส จัดอยู่ในประเภทลิกาส เช่น HMG-COA ซินเทส, ATP ซินเทส เช่น Succiny1-COA synthetase, Glutamine synthetase
Synthase vs Synthetase Comparison table
Watch This Video To Know About synthase vs synthetase <3
บทสรุป
- สารสังเคราะห์ไม่จำเป็นต้องใช้ NTP ในการทำงาน เนื่องจากสารสังเคราะห์เหล่านี้เร่งกระบวนการสังเคราะห์ที่ต้องการการย่อยสลายของนิวคลีโอไซด์ไตรฟอสเฟต (ไม่ใช่แค่ ATP)
- สหภาพระหว่างประเทศของ คณะกรรมการระบบการตั้งชื่อของชีวเคมีได้เปลี่ยนคำจำกัดความของซินเทสในทศวรรษที่ 1980 เพื่อให้ครอบคลุมเอ็นไซม์สังเคราะห์ทั้งหมด ไม่ว่าจะใช้ NTP หรือไม่ก็ตาม และซินเทเทสก็มีความหมายเหมือนกันกับลิเกส
- ลิเกสเป็นเอ็นไซม์ที่รวมโมเลกุลที่เล็กกว่าสองโมเลกุลเข้าด้วยกันโดยใช้ พลังงานจากการไฮโดรไลซิสของ NTP (โดยปกติจะผ่านการควบแน่นปฏิกิริยา).
- ซูโดริดีนซินเทส
- แฟตตีแอซิดซินเทส
- เซลลูโลสซินเทส (สร้าง UDP)
- เซลลูโลสซินเทส (สร้าง GDP)
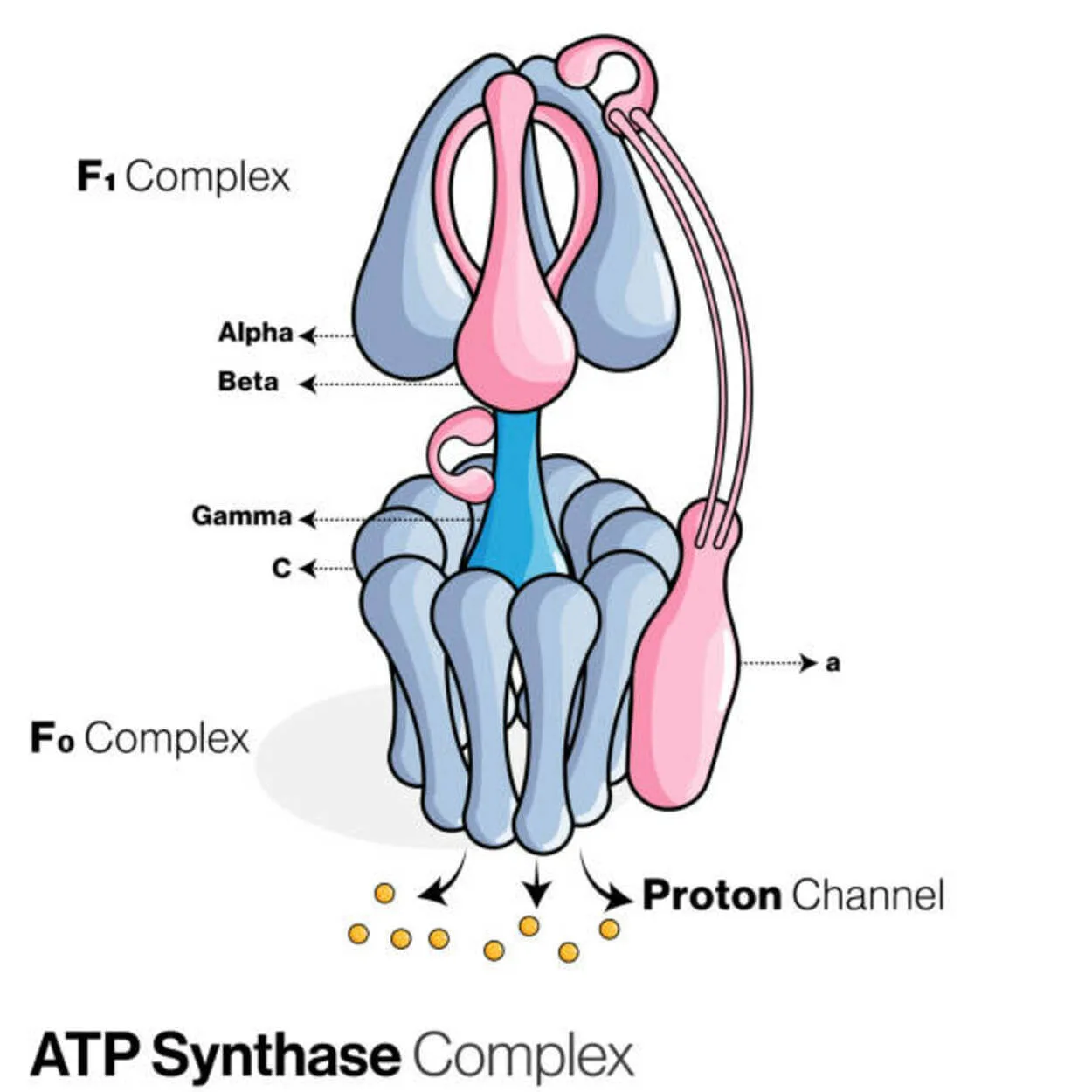
ATP Synthase
Adenosine diphosphate (ADP) และอนินทรีย์ฟอสเฟตถูกใช้เพื่อสร้างโมเลกุลกักเก็บพลังงาน adenosine triphosphate (ATP) โดยโปรตีนที่เรียกว่า ATP synthase (Pi)
มันถูกจัดประเภทเป็น ligase เนื่องจากมันแก้ไข ADP โดยสร้างลิงค์ P-O (พันธะฟอสโฟไดเอสเทอร์) อุปกรณ์ระดับโมเลกุลที่เรียกว่า ATP synthase
ในแง่พลังงาน การผลิต ATP จาก ADP และ Pi เป็นสิ่งที่ไม่พึงปรารถนา และโดยทั่วไปแล้วกระบวนการนี้จะไปในทางอื่น
การไล่ระดับความเข้มข้นของโปรตอน (H+) ผ่านเยื่อไมโทคอนเดรียด้านในในยูคารีโอตหรือพลาสมาเมมเบรนในแบคทีเรียจะขับเคลื่อนปฏิกิริยานี้ไปข้างหน้าโดยการควบรวมการสังเคราะห์ ATP ระหว่างการหายใจระดับเซลล์กับเกรเดียนต์
ในพืช ATP synthase ใช้โปรตอนเกรเดียนต์ที่เกิดขึ้นในไทลาคอยด์ลูเมนผ่านเยื่อหุ้มไทลาคอยด์และเข้าสู่คลอโรพลาสต์สโตรมาเพื่อผลิต ATP ระหว่างการสังเคราะห์ด้วยแสง
สำหรับ ATPase การสังเคราะห์เอทีพียูคาริโอตคือ F -ATPases ฟังก์ชันนั้น "ย้อนกลับ" ประเภทนี้จะถูกกล่าวถึงเป็นหลักในบทความนี้ หน่วยย่อย FO และ F1 ของ F-ATPase มีกลไกมอเตอร์หมุนที่ช่วยให้สังเคราะห์ ATP ได้
ซินเทสมีหลายประเภท
ซิเตรตซินเทส
เซลล์ที่มีชีวิตเกือบทั้งหมดมีเอนไซม์ซิเตรตซินเทสซึ่งทำหน้าที่เป็นเครื่องกระตุ้นหัวใจในขั้นตอนแรกของวงจรกรดซิตริก และกำหนด E.C. 2.3.3.1 (4.1.3.7 ก่อนหน้านี้) (หรือวงจรเครบ).
ซิเตรตซินเทสตั้งอยู่ในไมโตคอนเดรียลเมทริกซ์ของเซลล์ยูคารีโอต แม้ว่านิวเคลียส DNA จะเข้ารหัสมันซึ่งไม่ใช่ไมโตคอนเดรียลดีเอ็นเอ
มันถูกสร้างขึ้นในไซโตพลาสซึมโดยไรโบโซมของไซโตพลาสซึม และต่อมาถูกย้ายเข้าไปในเมทริกซ์ของไมโตคอนเดรีย
เครื่องหมายของเอนไซม์เชิงปริมาณทั่วไปสำหรับการมีอยู่ของไมโทคอนเดรียที่ไม่บุบสลายคือซิเตรตซินเทส กิจกรรมสูงสุดของซิเตรตซินเทสเผยให้เห็นว่ามีไมโทคอนเดรียจำนวนเท่าใดในกล้ามเนื้อโครงร่าง
การฝึกเป็นช่วงที่มีความเข้มข้นสูงมีศักยภาพในการเพิ่มกิจกรรมสูงสุดได้มากกว่าการฝึกความอดทนหรือการฝึกเป็นช่วงที่มีความเข้มข้นสูง
อะเซทิลโคเอนไซม์ A มีคาร์บอนอะซิเตตตกค้าง 2 โมเลกุลและโมเลกุล 4 โมเลกุล -carbon oxaloacetate กลั่นตัวเพื่อสร้างคาร์บอนซิเตรต 6 อะตอม ซึ่งผลิตโดยปฏิกิริยาการควบแน่นที่เร่งปฏิกิริยาโดยซิเตรตซินเทส
ทริปโตแฟนซินเทส
สองขั้นตอนสุดท้ายในการผลิตทริปโตเฟนคือ เร่งปฏิกิริยาโดยเอนไซม์ทริปโตเฟนซินเทสหรือที่เรียกว่าทริปโตเฟนซินเทเทส
Eubacteria, Archaebacteria, Protista, Fungi และ Plantae เป็นเจ้าภาพที่พบบ่อย อย่างไรก็ตาม Animalia ไม่มี โดยปกติแล้วจะปรากฏเป็น 2 2 tetramer
หน่วยย่อยเร่งปฏิกิริยาการเปลี่ยนกลับได้ของอินโดล-3-กลีเซอรอลฟอสเฟตไปเป็นอินโดลและกลีเซอรอลดีไฮด์-3-ฟอสเฟต (G3P) (IGP)
ในกระบวนการขึ้นกับไพริดอกซัลฟอสเฟต (PLP) หน่วยย่อยจะกระตุ้นการควบแน่นของอินโดลและซีรีนที่ผันกลับไม่ได้เพื่อสร้างทริปโตเฟน
ช่องทางที่ไม่ชอบน้ำภายในที่มีความยาว 25 อังสตรอมและอยู่ในเอนไซม์จะเชื่อมต่อแต่ละตำแหน่งที่ใช้งานกับตำแหน่งที่ใช้งานอยู่ติดกัน
สิ่งนี้ช่วยส่งเสริมช่องสัญญาณของซับสเตรต ซึ่งเป็นกลไกที่อินโดลผลิตขึ้นที่ไซต์ที่ใช้งานอยู่ แพร่กระจายโดยตรงไปยังไซต์อื่นที่ใช้งานอยู่ Tryptophan synthase ประกอบด้วย allosterically coupled active sites
ดูสิ่งนี้ด้วย: 2666 และ 3200 MHz RAM-อะไรคือความแตกต่าง? - ความแตกต่างทั้งหมดEubacteria, Archaebacteria, Protista, Fungi และ Plantae มักถูกค้นพบว่ามี tryptophan synthase รวมอยู่ด้วย มนุษย์และสัตว์อื่นขาดมัน
กรดอะมิโนที่จำเป็นสำหรับมนุษย์หนึ่งในเก้าชนิด ทริปโตเฟนเป็นหนึ่งในกรดอะมิโนมาตรฐานยี่สิบชนิด ทริปโตเฟนจึงจำเป็นต่ออาหารของมนุษย์
เป็นที่ทราบกันดีว่าทริปโตเฟนซินเทเทสสามารถใช้สารอะนาล็อกของอินโดล เช่น อินโดลที่มีฟลูออรีนหรือเมทิลเลต เป็นสารตั้งต้นเพื่อผลิตสารอะนาล็อกของทริปโตเฟนที่เทียบเท่ากัน
Pseudouridine
ตัวอักษรกรีก psi- ใช้เพื่อย่อ pseudouridine ซึ่งเป็นไอโซเมอร์ของ nucleoside uridine ซึ่ง uracil เชื่อมต่อกับอะตอมของคาร์บอนด้วยการเชื่อมโยงระหว่างคาร์บอนกับคาร์บอนแทนที่จะเป็นการเชื่อมต่อไกลโคซิดิกของไนโตรเจนกับคาร์บอน (Uracil บางครั้งเรียกว่า "pseudouracil" ในการจัดเรียงนี้)
RNA ที่แพร่หลายที่สุดการเปลี่ยนแปลงใน RNA ของเซลล์คือ pseudouridine RNA สามารถผ่านการเปลี่ยนแปลงทางเคมีที่ไม่ซ้ำกันมากกว่า 100 ครั้งระหว่างการถอดความและการสังเคราะห์
นอกเหนือจากนิวคลีโอไทด์ทั่วไปสี่ชนิดแล้ว สิ่งเหล่านี้อาจส่งผลต่อการแสดงออกของ RNA ภายหลังการถอดความ และมีหน้าที่หลายอย่างในเซลล์ รวมถึงการแปล RNA การโลคัลไลเซชัน และความเสถียร
หนึ่งในนั้นคือซูดูยูริดีน ซึ่งเป็นไอโซเมอร์ C5-ไกลโคไซด์ของยูริดีนที่มีพันธะ C-C ระหว่าง C1 ของน้ำตาลไรโบสกับ C5 ของยูราซิล แทนที่พันธะ C1-N1 ทั่วไปที่มีอยู่ในยูริดีน
มีความคล่องตัวในการหมุนเพิ่มเติมและความยืดหยุ่นของโครงสร้างเนื่องจากพันธะ CC นอกจากนี้ ตำแหน่ง N1 ของซูดูริดีนยังมีผู้บริจาคพันธะไฮโดรเจนเพิ่มเติม
ซูดูริดีนหรือที่เรียกว่า 5-ไรโบซิลลูราซิล เป็นองค์ประกอบที่คุ้นเคยแต่ลึกลับของอาร์เอ็นเอเชิงโครงสร้าง (การถ่ายโอน ไรโบโซม นิวเคลียร์ขนาดเล็ก (snRNA) และ นิวเคลียสขนาดเล็ก) เมื่อเร็ว ๆ นี้พบในการเข้ารหัส RNA เช่นกัน
เป็นชนิดแรกที่ถูกตรวจพบ เป็นที่แพร่หลายมากที่สุด และสามารถพบได้ในโดเมนวิวัฒนาการทั้งสามแห่งของสิ่งมีชีวิต ใน tRNA ของยีสต์ ซูโดยูริดีนสร้างนิวคลีโอไทด์ประมาณ 4%
ผ่านการสร้างพันธะไฮโดรเจนเพิ่มเติมกับน้ำ การเปลี่ยนแปลงเบสนี้สามารถทำให้ RNA เสถียรและเพิ่มการซ้อนเบสได้
จำนวนของ pseudouridines เพิ่มขึ้นตามความซับซ้อนของสิ่งมีชีวิต มี 11 pseudouridines ในrRNA ของ Escherichia coli, 30 ใน rRNA ของไซโตพลาสซึมของยีสต์, การเปลี่ยนแปลงหนึ่งครั้งใน 21S rRNA ของไมโทคอนเดรีย และประมาณ 100 ใน rRNA ของมนุษย์
มีการพิสูจน์แล้วว่า pseudouridine ใน rRNA และ tRNA ปรับจูนได้ และทำให้โครงสร้างระดับภูมิภาคมีเสถียรภาพและช่วยในการรักษาบทบาทในการถอดรหัส mRNA การประกอบไรโบโซม การประมวลผล และการแปล
มีการพิสูจน์แล้วว่า pseudouridine ใน snRNA ช่วยปรับปรุงอินเทอร์เฟซระหว่าง pre-mRNA และ spliceosomal RNA เพื่อช่วยควบคุมการประกบกัน
Fatty Acid Synthase
FASN ยีนในมนุษย์เข้ารหัสเอนไซม์ที่เรียกว่าการสังเคราะห์กรดไขมัน (FAS) โปรตีนหลายเอนไซม์ที่เรียกว่าการสังเคราะห์กรดไขมันจะกระตุ้นการสังเคราะห์กรดไขมัน
เป็นระบบเอนไซม์ทั้งหมด ไม่ใช่แค่เอนไซม์เดียว ประกอบด้วยโพลีเปปไทด์มัลติฟังก์ชั่นขนาด 272 kDa สองตัวที่เหมือนกันซึ่งถ่ายโอนสารตั้งต้นจากโดเมนการทำงานหนึ่งไปยังอีกโดเมนหนึ่ง
งานหลักของมันคือการใช้ NADPH เพื่อกระตุ้นการสร้างปาล์มมิเทต (C16:0, กรดไขมันอิ่มตัวสายยาว) จากอะเซทิล- และมาโลนิล-โคเอ
อะเซทิล-โคเอและมาโลนิล -CoA จะถูกเปลี่ยนเป็นกรดไขมันโดยลำดับของกระบวนการควบแน่น Claisen decarboxylative
ดูสิ่งนี้ด้วย: ที่เก็บข้อมูล SSD เทียบกับ eMMC (eMMC 32GB ดีกว่าไหม) – ความแตกต่างทั้งหมดหลังจากการยืดตัวในแต่ละรอบ คีโตรีดักเตส (KR) ดีไฮดราเทส (DH) และอีโนอิล รีดัคเตสจะทำงานตามลำดับเพื่อลดหมู่เบตาคีโตไปสู่ห่วงโซ่คาร์บอนที่อิ่มตัวอย่างสมบูรณ์(ER).
เมื่อสายโซ่ของกรดไขมันเติบโตจนมีความยาว 16 คาร์บอน จะถูกปลดปล่อยโดยการทำงานของไทโอเอสเทอเรส (TE) ซึ่งเชื่อมต่ออย่างโควาเลนต์กับกลุ่มเทียมฟอสโฟแพนธีอีนของโปรตีนตัวพาเอซิล (ACP) (กรดปาล์มิติก)
Cellulose Synthase (UDP-Forming)
เอนไซม์หลักที่รับผิดชอบในการผลิตเซลลูโลสคือ Cellulose Synthase (EC 2.4.1.12) ในรูปแบบ UDP-forming โดยทั่วไปเรียกว่า UDP-glucose: (1→4) 4-D-glucosyltransferase ของเอนไซม์สำหรับ D-glucan
GDP-glucose ถูกใช้โดยเอนไซม์ที่เกี่ยวข้องที่เรียกว่าเซลลูโลสซินเทส (GDP- ขึ้นรูป) (EC 2.4.1.29) ทั้งแบคทีเรียและพืชมีสมาชิกของเอนไซม์ตระกูลนี้
ส่วนประกอบของแบคทีเรียอาจเรียกอีกอย่างว่า BcsA (bacterial cellulose synthase) หรือ CelA ในขณะที่สมาชิกของพืชมักรู้จักกันในชื่อ CesA (cellulose synthase) หรือ CslA (คล้ายเซลลูโลสซินเทส) (เรียกง่ายๆ ว่า "เซลลูโลส") .
พืชได้รับ CesA อันเป็นผลมาจากเอนโดซิมไบโอซิสที่ก่อให้เกิดคลอโรพลาสต์ ตระกูลที่ 2 ของกลูโคซิลทรานสเฟอเรสรวมถึงตระกูลนี้ (GT2)
มวลชีวภาพส่วนใหญ่บนโลกผลิตขึ้นผ่านการสังเคราะห์ทางชีวภาพและการไฮโดรไลซิสโดยเอนไซม์ที่เรียกว่าไกลโคซิลทรานสเฟอเรส
เป็นที่ทราบกันดีว่าตระกูล CesA ของพืชประกอบด้วย วงศ์ย่อยเจ็ดวงศ์ และวงศ์ย่อยสาหร่ายพืชรวมมี 10 วงศ์
สัตว์กลุ่มเดียวที่มีเอนไซม์นี้คือurochordates ซึ่งได้รับจากการถ่ายโอนยีนแนวนอนเมื่อกว่า 530 ล้านปีก่อน
Cellulose Synthase (GDP-Forming)
เอนไซม์นี้เป็นสมาชิกของอนุวงศ์ย่อยของ hexosyltransferase ของ glycosyltransferases คลาสของเอนไซม์นี้เรียกตามชื่อวิทยาศาสตร์ว่า GDP-glucose:1,4-beta-D-glucan 4-beta-D-glucosyltransferase
ชื่ออื่นๆ ที่ใช้บ่อย ได้แก่ เซลลูโลสซินเทส (การสร้างกัวโนซีนไดฟอสเฟต) เซลลูโลสซินทีเทส และกัวโนซีนไดฟอสโฟกลูโคส-1,4-เบต้า-กลูแคนกลูโคซิลทรานสเฟอเรส เอนไซม์นี้มีส่วนร่วมในการเผาผลาญน้ำตาลซูโครสและแป้ง
Synthetase คืออะไร?
คำว่า "ซินทีเทส" ซึ่งบางครั้งเรียกว่า "ลิเกส" หมายถึงเอ็นไซม์กลุ่มใดกลุ่มหนึ่งที่มีประมาณ 50 ชนิดที่เร่งปฏิกิริยาเคมีอนุรักษ์พลังงานและไกล่เกลี่ยระหว่างเหตุการณ์การสลายตัวที่ใช้พลังงานและผลผลิต กระบวนการสังเคราะห์
การแยกพันธะฟอสเฟตที่มีพลังออก พวกมันสร้างพลังงานที่จำเป็นเพื่อกระตุ้นการรวมตัวของโมเลกุลสองโมเลกุล (ในหลายๆ กรณี โดยการเปลี่ยนอะดีโนซีนไตรฟอสเฟต [ATP] เป็นอะดีโนซีนไดฟอสเฟต [ADP] พร้อมกัน) .
ลิเกสที่เรียกว่ากรดอะมิโน-RNA ลิเกสเป็นตัวกระตุ้นการสร้างพันธะคาร์บอน-ออกซิเจนระหว่าง RNA ถ่ายโอนและกรดอะมิโน
เมื่อเอ็นไซม์บางชนิด เช่น เอไมด์ซินทีเทสและเปปไทด์ซินทีเทสทำงาน คาร์บอน-ไนโตรเจน (C-N)มีการสร้างพันธะขึ้น
Synthetase เรียกอีกอย่างว่า Ligase
ความแตกต่างระหว่าง Synthetase และ Synthase
Synthetase เป็นเอนไซม์ที่สามารถกระตุ้นการรวมตัวของ โมเลกุลขนาดใหญ่ 2 โมเลกุลโดยการสร้างพันธะเคมีใหม่ โดยปกติจะมีการไฮโดรไลซิสพร้อมกันของกลุ่มสารเคมีจี้ขนาดเล็กบนหนึ่งในโมเลกุลที่ใหญ่กว่า หรือสามารถกระตุ้นการเชื่อมโยงของสารประกอบ 2 ชนิด เช่น การรวมตัวกันของ C-O, C-S, C-N เป็นต้น
โดยทั่วไปแล้ว A ligase จะทำให้เกิดปฏิกิริยาต่อไปนี้:
- A-C + b = Ab + C
- A+D + B + C + D + E + F = Ab + cD
โดยที่กลุ่มเล็ก ๆ ที่ขึ้นต่อกันจะแสดงด้วยอักษรตัวพิมพ์เล็ก Ligase อาจซ่อมแซมการแตกตัวของสายเดี่ยวที่พัฒนาใน DNA สายคู่ระหว่างการจำลองแบบ รวมทั้งเชื่อมโยงชิ้นส่วนกรดนิวคลีอิกเสริมสองส่วน
ในทางกลับกัน ซินเทสเป็นเอนไซม์ที่กระตุ้นกระบวนการสังเคราะห์ทางชีวเคมี พวกมันรวมอยู่ในหมวดหมู่ของไลเอสตามการจัดหมวดหมู่หมายเลข EC
ระบบการตั้งชื่อ
โปรดจำไว้ว่า ระบบการตั้งชื่อทางชีววิทยาในขั้นต้นนั้นแยกความแตกต่างระหว่างการสังเคราะห์และการสังเคราะห์ ตามคำจำกัดความเดิม สารสังเคราะห์ใช้นิวคลีโอไซด์ไตรฟอสเฟต (เช่น ATP, GTP, CTP, TTP และ UTP) เป็นแหล่งพลังงานในขณะที่สังเคราะห์ไม่ได้
อย่างไรก็ตาม ตามที่คณะกรรมาธิการร่วมเกี่ยวกับการตั้งชื่อทางชีวเคมี (JCBN) กล่าวว่า "ซินเทส"