Mitä eroa on syntaasin ja syntetaasin välillä? (Faktat paljastuvat) - Kaikki erot

Sisällysluettelo
E.C. 6 Ligaasientsyymien luokkaan kuuluvat syntaasit ja syntetaasit. Ne osallistuvat synteettisiin reaktioihin ja katalysoivat kahden molekyylin yhdistymistä samalla kun ne katkaisevat ATP:n tai muun vastaavan trifosfaatin difosfaattilinkin.
Toisin kuin syntetaasit, jotka käyttävät ATP:tä energianlähteenä biologisten yhdisteiden synteesissä, syntaasit ovat mitä tahansa ligaaseja, jotka katalysoivat biologisten yhdisteiden synteesiä käyttämättä ATP:tä energianlähteenä.
Tässä artikkelissa kerrotaan tarkalleen, mitä eroa on syntaasin ja syntetaasin välillä.
Mikä on syntaasi?
Syntaasi on entsyymi, joka katalysoi synteesiprosessia biokemiassa. Muista, että biologisessa nimikkeistössä tehtiin alun perin ero syntetaasien ja syntaasien välillä.
Katso myös: Tuleeko kehossasi olemaan mitään eroa kuuden kuukauden kuluttua kuntosalilla? (Selvitä) - Kaikki erotAlkuperäisen määritelmän mukaan syntetaasit käyttävät nukleosiditrifosfaatteja (kuten ATP:tä, GTP:tä, CTP:tä, TTP:tä ja UTP:tä) energialähteenä, kun taas syntaasit eivät.
Joint Commission on Biochemical Nomenclature (JCBN) -järjestön mukaan "syntaasia" voidaan kuitenkin käyttää viittaamaan mihin tahansa entsyymiin, joka katalysoi synteesiä (riippumatta siitä, käyttääkö se nukleosiditrifosfaatteja), mutta "syntetaasia" tulisi käyttää vain viittaamaan "ligaasiin".
Seuraavassa on luettelo esimerkkejä erityyppisistä syntaaseista:
- ATP-syntaasi
- Sitraattisyntaasi
- Tryptofaanisyntaasi
- Pseudouridiinisyntaasi
- Rasvahapposyntaasi
- Selluloosasyntaasi (UDP:tä muodostava)
- Selluloosasyntaasi (GDP:tä muodostava)
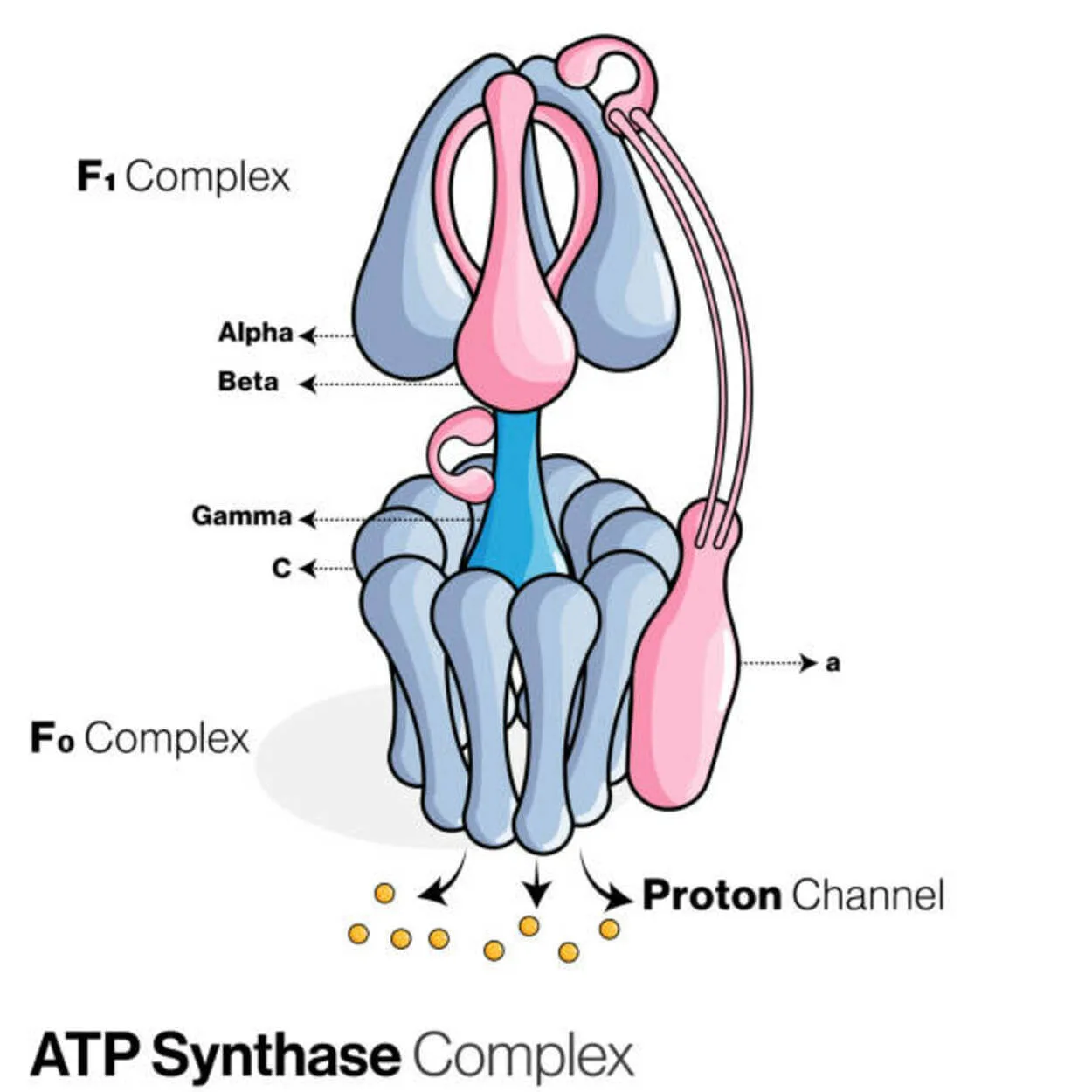
ATP-syntaasi
ATP-syntaasiksi (Pi) kutsuttu proteiini käyttää adenosiinidifosfaattia (ADP) ja epäorgaanista fosfaattia energiavarastomolekyylin adenosiinitrifosfaatin (ATP) muodostamiseen.
Se luokitellaan ligaasiksi, koska se muokkaa ADP:tä muodostamalla P-O-linkin (fosfodiesterisidoksen). Molekyylilaite, jota kutsutaan ATP-syntaasiksi.
Energian kannalta ATP:n tuottaminen ADP:stä ja Pi:stä ei ole toivottavaa, ja prosessi kulkisi yleensä päinvastoin.
Protonien (H+) pitoisuusgradientti eukaryoottien mitokondrioiden sisemmän kalvon tai bakteerien plasmakalvon yli vie tätä reaktiota eteenpäin kytkemällä ATP-synteesi soluhengityksen aikana gradienttiin.
Kasveissa ATP-syntaasi käyttää fotosynteesin aikana ATP:n tuottamiseen tylakoidilumeniin muodostunutta protonigradienttia tylakoidikalvon läpi ja kloroplastin stroomaan.
ATPaasin osalta eukaryoottiset ATP-syntaasit ovat F-ATPaaseja, jotka toimivat "käänteisesti". Tällaista käsitellään ensisijaisesti tässä artikkelissa. F-ATPaasin FO- ja F1-alayksiköillä on pyörivän moottorin mekanismi, joka mahdollistaa ATP-synteesin.
Syntaaseja on erityyppisiä
Sitraattisyntaasi
Lähes kaikissa elävissä soluissa on entsyymi sitraattisyntaasi, joka toimii tahdistimena sitruunahappokierron ensimmäisessä vaiheessa ja jota kutsutaan E.C. 2.3.3.1 (aiemmin 4.1.3.7). (tai Krebsin sykli).
Sitraattisyntaasi sijaitsee eukaryoottisolujen mitokondriomatriisissa, vaikka sitä koodaa ydin-DNA, ei mitokondrion DNA.
Se syntyy sytoplasmassa sytoplasman ribosomien toimesta ja siirretään sen jälkeen mitokondriomatriisiin.
Tyypillinen kvantitatiivinen entsyymimarkkeri ehjien mitokondrioiden olemassaololle on sitraattisyntaasi. Sitraattisyntaasin huippuaktiivisuus paljastaa, kuinka monta mitokondriaa luurankolihaksessa on.
Korkean intensiteetin intervalliharjoittelulla voidaan nostaa maksimaalista aktiivisuutta enemmän kuin kestävyysharjoittelulla tai korkean intensiteetin intervalliharjoittelulla.
Asetyylikoentsyymi A:ssa on kahden hiilen asetaattijäämä, ja neljän hiilen oksaloasetaattimolekyyli kondensoituu kuuden hiilen sitraatiksi, joka syntyy sitraattisyntaasin katalysoimassa kondensaatioreaktiossa.
Tryptofaanisyntaasi
Kaksi viimeistä vaihetta tryptofaanin tuotannossa katalysoi entsyymi tryptofaanisyntaasi, joka tunnetaan myös nimellä tryptofaanisyntetaasi.
Eubacteria, Archaebacteria, Protista, Fungi ja Plantae ovat usein sen isäntiä. Animalia ei kuitenkaan tunne sitä. Yleensä se esiintyy 2 2 -tetramerina.
Alayksiköt katalysoivat indoli-3-glyserolifosfaatin palautuvaa muuntumista indoliksi ja glyseraldehydi-3-fosfaatiksi (G3P) (IGP).
Pyridoksaalifosfaatista (PLP) riippuvaisessa prosessissa alayksiköt katalysoivat indolin ja seriinin irreversiibeliä kondensoitumista tryptofaaniksi.
Entsyymissä sijaitseva 25 angströmin pituinen sisäinen hydrofobinen kanava yhdistää kunkin aktiivisen alueen viereiseen aktiiviseen alueeseen.
Tämä edistää substraatin kanavoitumista, mekanismia, jonka avulla aktiivisissa kohdissa tuotettu indoli diffundoituu suoraan muihin aktiivisiin kohtiin. Tryptofaanisyntaasi sisältää allosterisesti kytkettyjä aktiivisia kohtia.
Eubakteereissa, arkebakteereissa, protistoissa, sienissä ja plantaeissa on usein havaittu tryptofaanisyntaasia. Ihmisiltä ja muilta eläimiltä se puuttuu.
Tryptofaani on yksi yhdeksästä ihmiselle välttämättömästä aminohaposta ja kuuluu kahdenkymmenen vakioaminohapon joukkoon. Tryptofaani on siis välttämätön ihmisen ruokavaliossa.
Tiedetään myös, että tryptofaanisyntetaasi voi käyttää indolianalogeja, kuten fluorattuja tai metyloituja indoleja, substraatteina tuottaakseen vastaavia tryptofaanianalogeja.
Pseudouridiini
Kreikankielistä kirjainta psi- käytetään lyhentämään pseudouridiinia, joka on uridiinin nukleosidin isomeeri, jossa urasiili on yhdistetty hiiliatomiin hiili-hiililinkillä eikä typpi-hiili-glykosidisella yhteydellä. (Uraciilia kutsutaan toisinaan "pseudourasiiliksi" tässä järjestelyssä.)
Solujen RNA:ssa yleisin RNA-muutos on pseudouridiini. RNA:ssa voi tapahtua yli 100 kemiallisesti ainutlaatuista muutosta transkription ja synteesin aikana.
Neljän tavanomaisen nukleotidin lisäksi nämä voivat mahdollisesti vaikuttaa RNA:n ilmentymiseen transkriptiota seuraten, ja niillä on useita tehtäviä solussa, kuten RNA:n translaatio, lokalisaatio ja stabiilisuus.
Yksi näistä on pseudouridiini, uridiinin C5-glykosidi-isomeeri, jossa riboosisokerin C1:n ja urasiilin C5:n välillä on C-C-sidos uridiinille tyypillisen C1-N1-sidoksen sijasta.
C-C-sidoksen ansiosta sillä on ylimääräistä rotaatioliikkuvuutta ja konformaatiojoustavuutta. Lisäksi pseudouridiinin N1-asemassa on ylimääräinen vetysidoksen luovuttaja.
Pseudouridiini, jota kutsutaan myös 5-ribosyyliurasiiliksi, on rakenteellisten RNA:iden (siirtorNA, ribosomaalinen RNA, pieni ydinrNA (snRNA) ja pieni nukleolaarinen RNA) tuttu mutta salaperäinen komponentti. Se on äskettäin löydetty myös koodaavasta RNA:sta.
Se havaittiin ensimmäisenä, se on yleisin, ja sitä esiintyy kaikilla kolmella evoluutioalueella. Hiivan tRNA:ssa pseudouridiini muodostaa noin 4 % nukleotideista.
Muodostamalla uusia vetysidoksia veden kanssa tämä emäsmuutos pystyy vakauttamaan RNA:ta ja parantamaan emästen pinoutumista.
Pseudouridiinien määrä kasvaa organismin monimutkaisuuden myötä. Escherichia coli -bakteerin rRNA:ssa on 11 pseudouridiinia, hiivan sytoplasmisessa rRNA:ssa 30, mitokondrion 21S-rRNA:ssa on yksi muutos ja ihmisen rRNA:ssa noin 100 pseudouridiinia.
On osoitettu, että rRNA:n ja tRNA:n sisältämä pseudouridiini hienosäätää ja vakauttaa alueellista rakennetta ja auttaa ylläpitämään niiden tehtäviä mRNA:n dekoodauksessa, ribosomien kokoamisessa, prosessoinnissa ja translaatiossa.
On osoitettu, että snRNA:n sisältämä pseudouridiini parantaa pre-mRNA:n ja spliceosomaalisen RNA:n välistä rajapintaa ja auttaa näin säätelemään splikointia.
Katso myös: Mitä eroa on "Donc" ja "Alors" välillä? (Yksityiskohtainen analyysi) - Kaikki erotRasvahapposyntaasi
Ihmisen FASN-geeni koodaa entsyymiä, joka tunnetaan nimellä rasvahapposyntaasi (FAS). Rasvahapposyntaasiksi kutsuttu moni-entsyyminen proteiini katalysoi rasvahappojen synteesiä.
Kyseessä on kokonainen entsymaattinen järjestelmä, ei vain yksi entsyymi, joka koostuu kahdesta identtisestä 272 kDa:n monitoimisesta polypeptidistä, jotka siirtävät substraatteja toiminnalliselta alueelta toiselle.
Sen ensisijainen tehtävä on katalysoida NADPH:n avulla palmitaatin (C16:0, pitkäketjuinen tyydyttynyt rasvahappo) muodostumista asetyyli- ja malonyyli-CoA:sta.
Asetyyli-CoA ja malonyyli-CoA muunnetaan rasvahapoiksi dekarboksylatiivisten Claisenin kondensaatioprosessien avulla.
Jokaisen pidennyskierroksen jälkeen ketoreduktaasi (KR), dehydraasi (DH) ja enoyylireduktaasi toimivat peräkkäin vähentääkseen beetaketoryhmän täysin tyydyttyneeksi hiiliketjuksi (ER).
Kun rasvahappoketju on kasvanut 16 hiilen pituiseksi, se vapautuu tioesteraasin (TE) vaikutuksesta, joka on kovalenttisesti kytkeytynyt asyylikuljettajaproteiinin (ACP) (palmitiinihappo) fosfopanteteiiniproteettiseen ryhmään.
Selluloosasyntaasi (UDP:tä muodostava)
Ensisijainen selluloosan tuottamisesta vastaava entsyymi on selluloosasyntaasi (EC 2.4.1.12) UDP-muodossaan. Sitä kutsutaan yleisesti D-glukaanin UDP-glukoosi: (1→4) entsyymologian 4-D-glukosyylitransferaasiksi.
BKT-glukoosia käyttää sukua oleva entsyymi nimeltä selluloosasyntaasi (BKT:tä muodostava) (EC 2.4.1.29). Sekä bakteereilla että kasveilla on tämän entsyymiperheen jäseniä.
Bakteerien jäsenet tunnetaan myös nimillä BcsA (bakteeriselluloosasyntaasi) tai CelA, kun taas kasvien jäsenet tunnetaan yleensä nimillä CesA (selluloosasyntaasi) tai spekulatiivinen CslA (selluloosasyntaasin kaltainen) (yksinkertaisesti "selluloosa").
Kasvit saivat CesA:n kloroplastin synnyttäneen endosymbioosin tuloksena. Glukosyylitransferaasien perheeseen 2 kuuluu tämä (GT2).
Suurin osa maapallon biomassasta tuotetaan biosynteesin ja hydrolyysin avulla glykosyylitransferaaseiksi kutsuttujen entsyymien avulla.
Kasvien CesA-superperheeseen tiedetään kuuluvan seitsemän alaperhettä, ja yhdistettyyn kasvi-levä-superperheeseen kuuluu 10 alaperhettä.
Ainoa eläinryhmä, jolla on tämä entsyymi, on urokordat, jotka saivat sen horisontaalisen geenisiirron kautta yli 530 miljoonaa vuotta sitten.
Selluloosasyntaasi (GDP:tä muodostava)
Tämä entsyymi kuuluu glykosyylitransferaasien heksosyylitransferaasi-alaperheeseen. Tähän entsyymiluokkaan viitataan sen tieteellisellä nimellä GDP-glukoosi:1,4-beta-D-glukaani 4-beta-D-glukosyylitransferaasi.
Muita usein käytettyjä nimityksiä ovat selluloosasyntaasi (guanosiinidifosfaattia muodostava), selluloosasyntetaasi ja guanosiinidifosfoglukoosi-1,4-beeta-glukaaniglukosyylitransferaasi. Tämä entsyymi osallistuu sakkaroosin ja tärkkelyksen aineenvaihduntaan.
Mikä on syntetaasi?
Termi "syntetaasi", joka tunnetaan joskus nimellä "ligaasi", viittaa mihin tahansa noin 50 entsyymin luokkaan, joka katalysoi energiaa säästäviä kemiallisia reaktioita ja välittää energiaa kuluttavien hajoamistapahtumien ja tuottavien synteettisten prosessien välillä.
Halkaisemalla energisen fosfaattisidoksen ne tuottavat tarvittavan energian kahden molekyylin yhdistymisen katalysoimiseksi (monissa tapauksissa muuntamalla adenosiinitrifosfaatin [ATP] samanaikaisesti adenosiinidifosfaatiksi [ADP]).
Aminohappo-RNA-ligaasiksi kutsuttu ligaasi katalysoi hiili-happisidoksen muodostumista siirto-RNA:n ja aminohapon välille.
Kun tietyt entsyymit, kuten amidisyntetaasit ja peptidisyntetaasit, ovat aktiivisia, syntyy hiili-typpi (C-N) -sidoksia.
Syntetaasi tunnetaan myös nimellä ligaasi
Syntetaasin ja syntetaasin välinen ero
Syntetaasi on entsyymi, joka voi katalysoida kahden suuren molekyylin yhdistämistä luomalla uuden kemiallisen sidoksen, tyypillisesti hydrolysoimalla samanaikaisesti pienen riippuvan kemiallisen ryhmän jossakin suuremmassa molekyylissä, tai se voi katalysoida kahden yhdisteen yhdistämistä, kuten C-O-, C-S-, C-N-, jne. yhdistämistä.
Ligaasi aiheuttaa tyypillisesti seuraavan reaktion:
- A-C + b = Ab + C
- A+D + B + C + D + E + F = Ab + cD = Ab + cD
Jos riippuvaiset, pienet ryhmittymät esitetään pienillä kirjaimilla. Ligaasi voi korjata kaksisäikeiseen DNA:han replikaation aikana syntyviä yksijuosteisia katkoksia sekä yhdistää kaksi toisiaan täydentävää nukleiinihappofragmenttia.
Toisaalta syntaasi on entsyymi, joka katalysoi synteesiprosessia biokemiassa. Ne kuuluvat EC-numeroluokituksen mukaan lyaasien luokkaan.
Nimikkeistö
On muistettava, että biologisessa nimikkeistössä tehtiin alun perin ero syntetaasien ja syntaasien välillä. Alkuperäisen määritelmän mukaan syntetaasit käyttävät nukleosiditrifosfaatteja (kuten ATP, GTP, CTP, TTP ja UTP) energianlähteenä, kun taas syntaasit eivät.
Joint Commission on Biochemical Nomenclature (JCBN) -järjestön mukaan "syntaasia" voidaan kuitenkin käyttää viittaamaan mihin tahansa entsyymiin, joka katalysoi synteesiä (riippumatta siitä, käyttääkö se nukleosiditrifosfaatteja), mutta "syntetaasia" tulisi käyttää vain viittaamaan "ligaasiin".
Ensisijainen ero syntaasin ja syntetaasin välillä on se, että syntetaasi on entsyymiperhe, joka voi luoda molekyylien välisiä sidoksia, kun taas syntaasi on entsyymi.
| Syntaasi | Syntetaasi |
| Katalysoi synteettistä prosessia ilman ATP:tä | Vaatii ATP:tä |
| Kuuluu maljakko- tai transferaasiluokitukseen | Kuuluu ligaasiluokitukseen |
| esim. HMG-COA-syntaasi, ATP-syntaasi. | esim. sukkini1-COA-syntetaasi, Glutamiini syntetaasi |
Vertailutaulukko syntaasi vs syntetaasi
Katso tämä video tietää syntaasi vs syntetaasi
Päätelmä
- Syntetaasit eivät tarvitse NTP:tä toimiakseen, koska ne katalysoivat synteettisiä prosesseja, jotka edellyttävät nukleosiditrifosfaattien hydrolyysiä (ei pelkästään ATP:tä).
- Kansainvälisen biokemian liiton nomenklatuurikomitea muutti syntaasin määritelmää 1980-luvulla siten, että se kattaa kaikki synteettiset entsyymit riippumatta siitä, käyttävätkö ne NTP:tä vai eivät, ja syntetaasista tuli synonyymi ligaasille.
- Ligaasi on entsyymi, joka yhdistää kaksi pienempää molekyyliä toisiinsa hyödyntämällä NTP:n hydrolyysistä saatavaa energiaa (yleensä kondensaatioreaktion avulla).