У чому різниця між синтазою та синтетазою (розкриті факти) - всі відмінності

Зміст
До класу лігазних ферментів E.C. 6 належать синтази і синтетази. Вони беруть участь у синтетичних реакціях і каталізують з'єднання двох молекул, одночасно розриваючи дифосфатний зв'язок в АТФ або іншому подібному трифосфаті.
На відміну від синтетаз, які використовують АТФ як джерело енергії під час синтезу біологічних сполук, синтази - це будь-які лігази, які каталізують синтез біологічних сполук без використання АТФ як джерела енергії.
У цій статті ви дізнаєтеся, в чому саме полягає різниця між синтетазою та синтетазою.
Що таке синтаза?
Синтаза - це фермент, який каталізує процес синтезу в біохімії. Майте на увазі, що в біологічній номенклатурі спочатку розрізняли синтетази та синтази.
Згідно з оригінальним визначенням, синтетази використовують нуклеозидтрифосфати (такі як АТФ, ГТФ, ЦТФ, ТТФ і УТФ) як джерело енергії, в той час як синтази - ні.
Проте, згідно з Об'єднаною комісією з біохімічної номенклатури (JCBN), "синтаза" може використовуватися для позначення будь-якого ферменту, який каталізує синтез (незалежно від того, чи використовує він нуклеозидтрифосфати), але "синтетаза" повинна використовуватися тільки для позначення "лігази".
Ось список прикладів різних типів синтаз:
- АТФ-синтаза
- Цитратсинтаза
- Триптофансинтаза
- Псевдоуридинсинтетаза
- Синтаза жирних кислот
- Целюлозосинтетаза (UDP-утворююча)
- Целюлозосинтетаза (ВВП-утворююча)
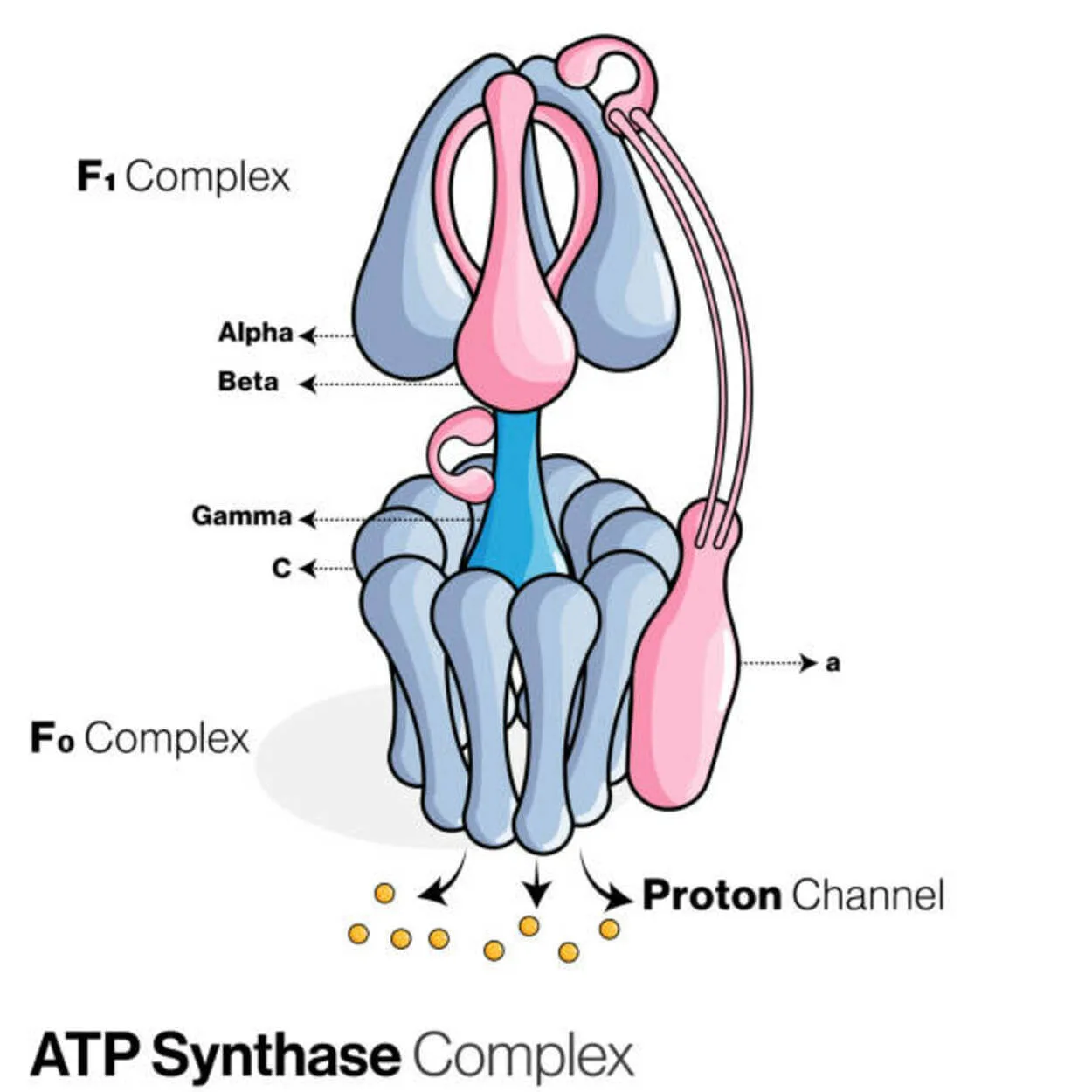
АТФ-синтаза
Аденозиндифосфат (АДФ) і неорганічний фосфат використовуються для створення молекули зберігання енергії аденозинтрифосфату (АТФ) білком, який називається АТФ-синтазою (Pi).
Його відносять до лігаз, оскільки він модифікує АДФ, утворюючи зв'язок P-O (фосфодіефірний зв'язок). Молекулярний пристрій, який називається АТФ-синтазою.
Дивіться також: Dupont Corian vs LG Hi-Macs: у чому відмінності - (Факти та відмінності) - Все про всеЗ енергетичної точки зору, виробництво АТФ з АДФ і Пі є небажаним, і процес, як правило, йде в зворотному напрямку.
Градієнт концентрації протонів (H+) через внутрішню мембрану мітохондрій в еукаріотів або плазматичну мембрану в бактерій рухає цю реакцію вперед, пов'язуючи синтез АТФ під час клітинного дихання з цим градієнтом.
У рослинах АТФ-синтаза використовує градієнт протонів, що утворюється в просвіті тилакоїда, через мембрану тилакоїда і в строму хлоропласта для виробництва АТФ під час фотосинтезу.
Що стосується АТФ-ази, то еукаріотичні АТФ-синтази - це F-АТФази, які функціонують "у зворотному напрямку". Саме цей тип розглядається в цій статті. Субодиниці FO і F1 F-АТФази мають обертальний руховий механізм, який забезпечує синтез АТФ.
Існують різні типи синтаз
Цитратсинтаза
Майже всі живі клітини містять фермент цитратсинтазу, який виконує роль пейсмейкера на першому етапі циклу лимонної кислоти і позначений К.К. 2.3.3.1 (раніше 4.1.3.7) (або цикл Кребса).
Цитратсинтаза знаходиться в мітохондріальному матриксі еукаріотичних клітин, хоча кодує її ядерна ДНК, а не мітохондріальна.
Він створюється в цитоплазмі за допомогою цитоплазматичних рибосом і згодом переміщується в мітохондріальний матрикс.
Типовим кількісним ферментним маркером існування інтактних мітохондрій є цитратсинтаза. Пікова активність цитратсинтази показує, скільки мітохондрій присутні в скелетному м'язі.
Високоінтенсивне інтервальне тренування здатне підвищити максимальну активність більше, ніж тренування на витривалість або високоінтенсивне інтервальне тренування.
Ацетилкоензим А має двовуглецевий ацетатний залишок, а молекула чотиривуглецевого оксалоацетату конденсується з утворенням шестивуглецевого цитрату, який утворюється в результаті реакції конденсації, каталізованої цитратсинтазою.
Синтаза триптофану
Останні два етапи виробництва триптофану каталізуються ферментом триптофансинтетазою, також відомим як триптофансинтетаза.
Еубактерії, архебактерії, протеї, гриби та рослини є його частими господарями. Однак у тварин його немає. Зазвичай він представлений у вигляді тетрамеру 2 2.
Субодиниці каталізують оборотне перетворення індол-3-гліцеролфосфату в індол і гліцеральдегід-3-фосфат (G3P) (IGP).
У процесі, залежному від піридоксальфосфату (PLP), субодиниці каталізують незворотну конденсацію індолу та серину з утворенням триптофану.
Внутрішній гідрофобний канал довжиною 25 ангстрем, розташований у ферменті, з'єднує кожну активну ділянку з сусідньою активною ділянкою.
Це сприяє субстратному каналуванню - механізму, за допомогою якого індол, що утворюється на активних ділянках, дифундує безпосередньо до інших активних ділянок. Триптофансинтаза містить алостерично з'єднані активні ділянки.
Еубактерії, архебактерії, протисти, гриби та рослини часто містять триптофансинтазу. Люди та інші тварини не мають її.
Одна з дев'яти необхідних для людини амінокислот, триптофан є однією з двадцяти стандартних амінокислот. Тому триптофан має важливе значення для людського раціону.
Також відомо, що триптофансинтетаза може використовувати аналоги індолів, такі як фторовані або метильовані індоли, як субстрати для виробництва еквівалентних аналогів триптофану.
Псевдоуридин
Грецька літера psi- використовується для скорочення псевдоуридину, ізомеру нуклеозиду уридину, в якому урацил приєднаний до атома вуглецю не глікозидним зв'язком азот-вуглець, а зв'язком вуглець-вуглець (урацил іноді називають "псевдоурацилом" у такому розташуванні).
Найпоширенішою зміною клітинної РНК є псевдоуридинова. РНК може зазнавати понад 100 хімічно унікальних змін під час транскрипції та синтезу.
Дивіться також: У чому різниця між JTAC і TACP (відмінність) - всі відмінностіНа додаток до чотирьох звичайних нуклеотидів, вони можуть потенційно впливати на експресію РНК посттранскрипційно і виконувати ряд функцій у клітині, включаючи трансляцію, локалізацію та стабільність РНК.
Одним з них є псевдоуридин, C5-глікозидний ізомер уридину з C-C зв'язком між C1 цукру рибози і C5 урацилу замість типового C1-N1 зв'язку, присутнього в уридині.
Він має додаткову обертальну рухливість і конформаційну гнучкість завдяки зв'язку С-С. Крім того, положення N1 псевдоуридину має додатковий донор водневого зв'язку.
Псевдоуридин, також відомий як 5-рибозилурацил, є знайомим, але загадковим компонентом структурних РНК (транспортних, рибосомальних, малих ядерних (snRNA) та малих нуклеолярних). Нещодавно його було виявлено також у кодуючих РНК.
Він був виявлений першим, є найпоширенішим і зустрічається у всіх трьох еволюційних сферах життя. У тРНК дріжджів псевдоуридин становить близько 4% нуклеотидів
Завдяки утворенню додаткових водневих зв'язків з водою ця зміна основи здатна стабілізувати РНК і посилити укладання основ.
Кількість псевдоуридинів зростає зі складністю організму. 11 псевдоуридинів є в рРНК кишкової палички, 30 - в цитоплазматичній рРНК дріжджів, одна зміна в мітохондріальній 21S рРНК і приблизно 100 - в рРНК людини.
Було продемонстровано, що псевдоуридин в рРНК і тРНК тонко налаштовує і стабілізує регіональну структуру і сприяє підтримці їхньої ролі в декодуванні мРНК, збірці рибосом, процесингу і трансляції.
Було продемонстровано, що псевдоуридин у складі іРНК покращує інтерфейс між пре-мРНК та сплайсосомальною РНК, допомагаючи регулювати сплайсинг.
Синтаза жирних кислот
Ген FASN у людини кодує фермент, відомий як синтаза жирних кислот (FAS). Мультиферментний білок під назвою синтаза жирних кислот каталізує синтез жирних кислот.
Це ціла ферментативна система, а не просто один фермент, що складається з двох ідентичних багатофункціональних поліпептидів по 272 кДа, які переносять субстрати з одного функціонального домену в інший.
Його основне завдання полягає у використанні НАДФН для каталізації утворення пальмітату (C16:0, довголанцюгової насиченої жирної кислоти) з ацетил- і малоніл-КоА.
Ацетил-КоА і малоніл-КоА перетворюються на жирні кислоти через послідовність декарбоксилюючих процесів конденсації Клейзена.
Після кожного раунду елонгації кеторедуктаза (KR), дегідратаза (DH) та еноілредуктаза працюють послідовно, щоб зменшити бета-кетогрупу до повністю насиченого вуглецевого ланцюга (ER).
Коли ланцюг жирної кислоти виростає до довжини 16 вуглеців, вона вивільняється під дією тіоестерази (ТЕ), яка ковалентно з'єднується з фосфопантеїновою простетичною групою ацильного білка-переносника (АБП) (пальмітинової кислоти).
Целюлозосинтетаза (UDP-утворююча)
Основним ферментом, відповідальним за утворення целюлози, є целюлозосинтаза (EC 2.4.1.12) в її УДФ-формі. Її зазвичай називають УДФ-глюкозою: (1→4) ензимологічна 4-D-глюкозилтрансфераза для D-глюкану.
Глюкоза використовується спорідненим ферментом, який називається целюлозосинтетаза (GDP-утворююча) (EC 2.4.1.29). Представники цієї родини ферментів є як у бактерій, так і у рослин.
Бактеріальні члени також можуть бути відомі як BcsA (бактеріальна целюлозосинтетаза) або CelA, тоді як рослинні члени зазвичай відомі як CesA (целюлозосинтетаза) або спекулятивна назва CslA (целюлозосинтетазоподібна) (просто "целюлоза").
CesA була придбана рослинами в результаті ендосимбіозу, який дав початок хлоропласту. Сімейство 2 глюкозилтрансфераз включає в себе цю ферментну систему (GT2).
Більшість біомаси на Землі виробляється шляхом біосинтезу та гідролізу за допомогою ферментів, які називаються глікозилтрансферазами.
Відомо, що надродина рослин CesA містить сім підродин, а об'єднана надродина рослинно-водоростева - 10.
Єдиною групою тварин, яка володіє цим ферментом, є урохорди, які отримали його шляхом горизонтального перенесення генів понад 530 мільйонів років тому.
Целюлозосинтетаза (ВВП-утворююча)
Цей фермент є членом підродини гексозилтрансфераз глікозилтрансфераз. Цей клас ферментів має наукову назву GDP-глюкоза:1,4-бета-D-глюкан 4-бета-D-глюкозилтрансфераза.
Інші назви, які часто використовуються: целюлозосинтетаза (гуанозиндифосфатоутворююча), целюлозосинтетаза, гуанозиндифосфоглюкозо-1,4-бета-глюканглюкозилтрансфераза. Цей фермент бере участь у метаболізмі сахарози і крохмалю.
Що таке синтетаза?
Термін "синтетаза", іноді відомий як "лігаза", відноситься до будь-якого класу з приблизно 50 ферментів, які каталізують хімічні енергозберігаючі реакції і є посередниками між енерговитратними процесами розщеплення і продуктивними синтетичними процесами.
Розщеплюючи енергійний фосфатний зв'язок, вони генерують необхідну енергію для каталізації з'єднання двох молекул (у багатьох випадках шляхом одночасного перетворення аденозинтрифосфату [АТФ] в аденозиндифосфат [АДФ]).
Лігаза, відома як амінокислотно-РНК-лігаза, каталізує утворення вуглецево-кисневого зв'язку між транспортною РНК та амінокислотою.
Коли певні ферменти, такі як амідосинтетази та пептидсинтетази, активні, утворюються вуглець-азотні (C-N) зв'язки.
Синтетаза також відома як лігаза
Різниця між синтетазою та синтазою
Синтетаза - це фермент, який може каталізувати з'єднання двох великих молекул шляхом створення нового хімічного зв'язку, як правило, з одночасним гідролізом невеликої підвішеної хімічної групи на одній з великих молекул, або може каталізувати з'єднання двох сполук, наприклад, з'єднання C-O, C-S, C-N тощо.
Лігаза зазвичай викликає наступну реакцію:
- A-C + b = Ab + C
- A + D + B + C + D + E + F = Ab + cD
Де залежні крихітні угруповання позначені малими літерами. Лігаза може відновлювати одноланцюгові розриви, що виникають у дволанцюговій ДНК під час реплікації, а також з'єднувати два комплементарні фрагменти нуклеїнової кислоти.
З іншого боку, синтаза - це фермент, який каталізує процес синтезу в біохімії. Вони входять до категорії ліаз відповідно до класифікації EC-номерів.
Номенклатура
Майте на увазі, що в біологічній номенклатурі спочатку розрізняли синтетази і синтази. Відповідно до початкового визначення, синтетази використовують нуклеозидтрифосфати (такі як АТФ, ГТФ, ЦТФ, ТТФ і УТФ) як джерело енергії, в той час як синтази - ні.
Проте, згідно з Об'єднаною комісією з біохімічної номенклатури (JCBN), "синтаза" може використовуватися для позначення будь-якого ферменту, який каталізує синтез (незалежно від того, чи використовує він нуклеозидтрифосфати), але "синтетаза" повинна використовуватися тільки для позначення "лігази".
Основна відмінність між синтетазою і синтетазою полягає в тому, що синтетаза - це сімейство ферментів, які можуть створювати зв'язки між молекулами, в той час як синтетаза - це фермент.
| Синтез | Синтетаза |
| Каталізує синтетичний процес без АТФ | Потребує АТФ |
| Підпадає під класифікацію ваз або трансформерів | Підпадає під класифікацію лігаз |
| наприклад, HMG-COA-синтаза, АТФ-синтаза | наприклад, сукциніл1-COA синтетаза, глютамін синтетаза |
Порівняльна таблиця "Синтетаза vs Синтетаза
Подивіться це відео, щоб дізнатися про різницю між синтазою та синтетазою
Висновок
- Синтетази не потребують NTP для функціонування, оскільки вони каталізують синтетичні процеси, які вимагають гідролізу нуклеозидтрифосфатів (а не просто АТФ).
- У 1980-х роках Номенклатурний комітет Міжнародного союзу біохімії змінив визначення синтази, включивши до нього всі синтетичні ферменти, незалежно від того, використовують вони NTP чи ні, і синтетаза стала синонімом лігази.
- Лігаза - це фермент, який з'єднує дві менші молекули разом, використовуючи енергію від гідролізу NTP (зазвичай через реакцію конденсації).