Kāda ir atšķirība starp sintēzi un sintēzi? (Fakti atklāti) - Visas atšķirības

Satura rādītājs
E.C. 6 ligāžu enzīmu klasē ietilpst sintāzes un sintetāzes. Tās piedalās sintētiskajās reakcijās un katalizē divu molekulu savienošanu, vienlaikus pārraujot difosfāta savienojumu ATP vai citā salīdzināmā trifosfātā.
Atšķirībā no sintetāzēm, kas bioloģisko savienojumu sintēzes laikā kā enerģijas avotu izmanto ATP, sintāzes ir jebkuras ligāzes, kas katalizē bioloģisko savienojumu sintēzi, neizmantojot ATP kā enerģijas avotu.
Šajā rakstā uzzināsiet, kāda ir atšķirība starp sintēzi un sintēzi.
Kas ir sintēze?
Sintāze ir enzīms, kas bioķīmijā katalizē sintēzes procesu. Jāatceras, ka bioloģiskajā nomenklatūrā sākotnēji tika nošķirtas sintēzes un sintāzes.
Skatīt arī: Cik ilgi princis palika nolādēts kā zvērs? Kāda ir vecuma atšķirība starp Bellu un Zvēru? (Detalizēti) - Visas atšķirībasSaskaņā ar sākotnējo definīciju sintetāzes kā enerģijas avotu izmanto nukleozīdu trifosfātus (piemēram, ATP, GTP, CTP, TTP un UTP), bet sintetāzes to neizmanto.
Tomēr saskaņā ar Apvienotās bioķīmiskās nomenklatūras komisijas (Joint Commission on Biochemical Nomenclature - JCBN) atzinumu terminu "sintēze" var lietot, lai apzīmētu jebkuru fermentu, kas katalizē sintēzi (neatkarīgi no tā, vai tajā tiek izmantoti nukleozīdu trifosfāti), bet terminu "sintēze" drīkst lietot tikai attiecībā uz "ligāzi".
Šeit ir saraksts ar dažādu sintēzes veidu piemēriem:
- ATP sintēze
- Citrāta sintēze
- Triptofāna sintēze
- Pseidouridīna sintēze
- Taukskābju sintēze
- Celulozes sintēze (UDP veidojošā)
- Celulozes sintēze (veidojošā GDP)
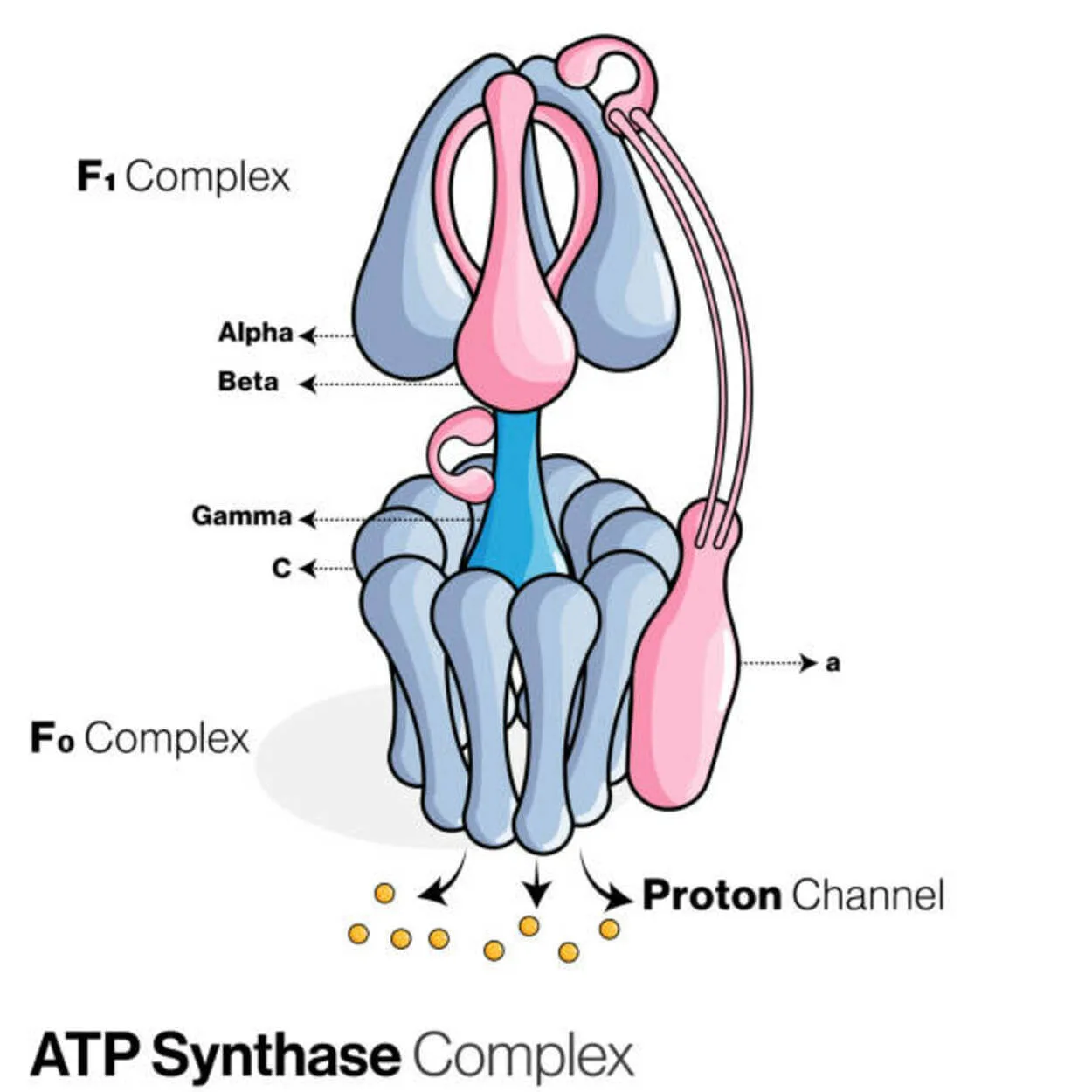
ATP sintēze
Adenozīna difosfātu (ADP) un neorganisko fosfātu izmanto, lai, izmantojot olbaltumvielu, ko sauc par ATP sintāzi (Pi), izveidotu enerģijas uzkrāšanas molekulu adenozīna trifosfātu (ATP).
Tā tiek klasificēta kā ligāze, jo tā modificē ADP, veidojot P-O saiti (fosfodiestera saiti). Molekulāro ierīci sauc par ATP sintēzi.
Enerģijas ziņā ATP ražošana no ADP un Pi ir nevēlama, un process parasti notiek pretējā virzienā.
Protonu (H+) koncentrācijas gradients pāri iekšējai mitohondriju membrānai eikariontos vai plazmas membrānai baktērijās virza šo reakciju uz priekšu, saistot ATP sintēzi šūnu elpošanas laikā ar šo gradientu.
Augos ATP sintēze izmanto protonu gradientu, kas veidojas tilakoīdu lūmenā, lai fotosintēzes laikā ražotu ATP, šķērsojot tilakoīdu membrānu un iekļūstot hloroplastu stromā.
ATPāzei eikariotu ATP sintēzes ir F-ATPāzes, kas darbojas "otrādi." Šis veids galvenokārt tiek aplūkots šajā rakstā. F-ATPāzes FO un F1 apakšvienībām ir rotācijas motora mehānisms, kas nodrošina ATP sintēzi.
Ir dažādi sintāzes veidi
Citrāta sintēze
Gandrīz visās dzīvās šūnās ir enzīms citrātsintāze, kas kalpo kā citronskābes cikla pirmā posma stimulators un tiek apzīmēts ar E.C. 2.3.3.3.1. (iepriekš 4.1.3.3.7.) (jeb Krebsa cikls).
Citrātsintetāze atrodas eikariotisko šūnu mitohondriālajā matricā, lai gan kodē kodola DNS, nevis mitohondriālā DNS.
To citoplazmā veido citoplazmas ribosomas un pēc tam pārvieto uz mitohondriju matricu.
Tipisks kvantitatīvs enzīmu marķieris, kas norāda uz neskartu mitohondriju esamību, ir citrātsintetāze. Citrātsintetāzes maksimālā aktivitāte parāda, cik daudz mitohondriju ir skeleta muskuļos.
Augstas intensitātes intervālu treniņiem ir potenciāls palielināt maksimālo aktivitāti vairāk nekā izturības treniņiem vai augstas intensitātes intervālu treniņiem.
Acetilkoenzīma A molekulā ir divu ogļhidrātu acetāta atlikums, un četru ogļhidrātu oksalacetāta molekula kondensējas, veidojot sešu ogļhidrātu citrātu, kas rodas kondensācijas reakcijā, ko katalizē citrātsintetāze.
Triptofāna sintēze
Pēdējos divus triptofāna ražošanas posmus katalizē ferments triptofāna sintēze, ko dēvē arī par triptofāna sintetāzi.
Eubacteria, Archaebacteria, Protista, Fungi un Plantae ir tās biežākie saimnieki. Tomēr Animalia tās nav. Parasti tā parādās kā 2 2 tetramērs.
Apakšvienības katalizē atgriezenisku indol-3-glicerīna fosfāta pārveidi par indolu un gliceraldehīda-3-fosfātu (G3P) (IGP).
Piridoksalfosfāta (PLP) atkarīgā procesā apakšvienības katalizē neatgriezenisku indola un serīna kondensāciju, lai iegūtu triptofānu.
Iekšējais hidrofobais kanāls, kas ir 25 angstroms garš un atrodas fermentā, savieno katru aktīvo vietu ar blakus esošo aktīvo vietu.
Tas veicina substrāta kanalizāciju - mehānismu, ar kura palīdzību aktīvajās vietās radies indols difundē tieši uz citām aktīvajām vietām. Triptofāna sintēzē ir alosteriski saistītas aktīvās vietas.
Eubacteria, Archaebacteria, Protista, Fungi un Plantae bieži vien satur triptofāna sintāzi. Cilvēkiem un citiem dzīvniekiem tās nav.
Triptofāns ir viena no deviņām cilvēkam nepieciešamajām aminoskābēm un viena no divdesmit standarta aminoskābēm. Tāpēc triptofāns ir būtisks cilvēka uzturā.
Ir arī zināms, ka triptofāna sintetāze var izmantot indola analogus, piemēram, fluorētus vai metilētus indolus, kā substrātus, lai iegūtu līdzvērtīgus triptofāna analogus.
Pseidouridīns
Grieķu burtu psi- lieto, lai saīsināti apzīmētu pseidouridīnu, nukleozīda uridīna izomēru, kurā uracils ir savienots ar oglekļa atomu ar oglekļa-oglekļa savienojumu, nevis slāpekļa-oglekļa glikozīdisku savienojumu (Uracils dažkārt tiek saukts par "pseidouracilu" šādā izkārtojumā).
Visizplatītākā RNS izmaiņa šūnu RNS ir pseidouridīns. Transkripcijas un sintēzes laikā RNS var notikt vairāk nekā 100 ķīmiski unikālu izmaiņu.
Papildus četriem parastajiem nukleotīdiem tie potenciāli var ietekmēt RNS ekspresiju pēc transkripcijas, un tiem ir vairākas funkcijas šūnā, tostarp RNS translācija, lokalizācija un stabilitāte.
Viens no tiem ir pseidouridīns, uridīna C5-glikozīda izomērs ar C-C saiti starp ribozes cukura C1 un uracila C5, kas aizvieto tipisko C1-N1 saiti, kura ir raksturīga uridīnam.
Pseidouridīna N1 pozīcijā ir papildu rotācijas kustība un konformācijas elastība, jo tajā ir C-C saite. Turklāt pseidouridīna N1 pozīcijā ir papildu ūdeņraža saites donors.
Pseidouridīns, saukts arī par 5-ribosiluracilu, ir pazīstama, bet noslēpumaina strukturālo RNS (transfēra, ribosomālo, mazo kodolu (snRNA) un mazo nukleāro RNS) sastāvdaļa. Nesen tas tika atklāts arī kodējošajās RNS.
Tas tika atklāts pirmais, ir visizplatītākais un sastopams visās trijās dzīvības evolūcijas jomās. Rauga tRNA pseidouridīns veido aptuveni 4 % no nukleotīdiem.
Veidojoties papildu ūdeņraža saitēm ar ūdeni, šī bāzes maiņa spēj stabilizēt RNS un uzlabot bāzes sakopošanu.
Pseidouridīnu skaits pieaug līdz ar organisma sarežģītību. 11 pseidouridīni ir Escherichia coli rRNS, 30 - rauga citoplazmas rRNS, viena izmaiņa mitohondriālajā 21S rRNS un aptuveni 100 - cilvēka rRNS.
Ir pierādīts, ka pseidouridīns rRNS un tRNS precizē un stabilizē reģionālo struktūru un palīdz saglabāt to lomu mRNS dekodēšanā, ribosomu montāžā, apstrādē un translācijā.
Ir pierādīts, ka pseidouridīns snRNA sastāvā uzlabo saskarni starp pre-mRNS un splicosomālo RNS, lai palīdzētu regulēt splicēšanu.
Taukskābju sintēze
FASN gēns cilvēkiem kodē enzīmu, kas pazīstams kā taukskābju sintāze (FAS). Daudzenzīmu proteīns, ko sauc par taukskābju sintāzi, katalizē taukskābju sintēzi.
Tā ir vesela fermentu sistēma, nevis tikai viens enzīms, kas sastāv no diviem identiskiem 272 kDa daudzfunkcionāliem polipeptīdiem, kuri pārnes substrātus no viena funkcionālā apgabala uz otru.
Tās galvenais uzdevums ir izmantot NADPH, lai katalizētu palmitāta (C16:0, garas ķēdes piesātinātās taukskābes) veidošanos no acetil- un malonil-CoA.
Acetil-CoA un malonil-CoA tiek pārvērsti taukskābēs, izmantojot dekarboksilātu Claisen kondensācijas procesu secību.
Pēc katras pagarināšanas kārtas secīgi darbojas ketoreduktāze (KR), dehidratāze (DH) un enoilreduktāze, lai samazinātu beta keto grupu līdz pilnīgi piesātinātai oglekļa ķēdei (ER).
Kad taukskābju ķēde ir palielinājusies līdz 16 ogļhidrātu garumam, to atbrīvo, iedarbojoties tioesterāzei (TE), kas kovalentizēti savienojas ar acilnesēja proteīna (ACP) (palmitīnskābe) fosfopanteheīna prostētisko grupu.
Celulozes sintēze (UDP-formējošā)
Galvenais enzīms, kas atbild par celulozes ražošanu, ir celulozes sintāzes (EC 2.4.1.12) UDP-formas forma. Parasti to dēvē par UDP-glikozes: (1→4) Enzimoloģijas 4-D-glikoziltransferāzi D-glikāna gadījumā.
GDP-glikozi izmanto radniecīgs enzīms, ko sauc par celulozes sintēzi (GDP-formējošo) (EC 2.4.1.29). Gan baktērijām, gan augiem ir šīs saimes enzīmu pārstāvji.
Baktēriju locekļus var dēvēt arī par BcsA (baktēriju celulozes sintēzi) vai CelA, bet augu locekļus parasti dēvē par CesA (celulozes sintēzi) vai spekulatīvo CslA (celulozes sintēzei līdzīgo) (vienkārši "celulozi").
CesA augi ieguva endosimbiozes rezultātā, kuras rezultātā radās hloroplasts. 2. glikoziltransferāžu saimē ietilpst šī (GT2).
Lielākā daļa biomasas uz Zemes tiek saražota biosintēzes un hidrolīzes ceļā, ko veic enzīmi, ko sauc par glikoziltransferāzēm.
Ir zināms, ka augu CesA superģimenei ir septiņas apakšģimenes, bet kombinētajai augu un aļģu superģimenei - 10.
Vienīgā dzīvnieku grupa, kurai ir šis ferments, ir urohordāti, kas to ieguva horizontālās gēnu pārneses ceļā pirms vairāk nekā 530 miljoniem gadu.
Celulozes sintēze (veidojošā GDP)
Šis enzīms ir glikoziltransferāžu apakšģimenes heksoziltransferāžu grupas loceklis. Šo enzīmu klasi apzīmē ar zinātnisko nosaukumu GDP-glikozes:1,4-beta-D-glikāna 4-beta-D-glikoziltransferāze.
Citi bieži lietotie nosaukumi ir celulozes sintetāze (veidojošā gvanozīna difosfātu), celulozes sintetāze un gvanozīna difosfoglikozes-1,4-beta-glikāna glikoziltransferāze. Šis ferments piedalās saharozes un cietes metabolismā.
Kas ir sintēze?
Termins "sintetāze", dažkārt saukts arī par "ligāzi", attiecas uz jebkuru no aptuveni 50 enzīmu klases, kas katalizē ķīmiskas enerģiju taupošas reakcijas un ir starpnieks starp enerģiju patērējošiem sadalīšanās procesiem un produktīviem sintēzes procesiem.
Sašķeļot enerģētisko fosfāta saiti, tie rada nepieciešamo enerģiju, lai katalizētu divu molekulu savienošanu (daudzos gadījumos vienlaicīgi pārveidojot adenozīna trifosfātu [ATP] par adenozīna difosfātu [ADP]).
Par aminoskābes-RNA ligāzi sauc aminoskābes-RNA ligāzi, kas katalizē oglekļa-skābekļa saites veidošanos starp transfēra RNS un aminoskābi.
Kad daži enzīmi, piemēram, amīdu sintēzes un peptīdu sintēzes, ir aktīvi, veidojas oglekļa un slāpekļa (C-N) saites.
Sintetāzi sauc arī par ligāzi
Atšķirība starp sintēzi un sintēzi
Sintetāze ir enzīms, kas var katalizēt divu lielu molekulu savienošanu, izveidojot jaunu ķīmisko saiti, parasti vienlaicīgi hidrolizējot nelielu ķīmisko grupu, kas atrodas uz vienas no lielākajām molekulām, vai arī var katalizēt divu savienojumu savienošanu, piemēram, C-O, C-S, C-N u. c. savienošanu.
Līgaze parasti izraisa šādu reakciju:
Skatīt arī: Kāda ir atšķirība starp IPS monitoru un LED monitoru (detalizēts salīdzinājums) - visas atšķirības- A-C + b = Ab + C
- A+D + B + C + D + E + F = Ab + cD
Ja atkarīgās, sīkās grupas apzīmētas ar mazajiem burtiem. Ligaze var labot vienas virknes pārrāvumus, kas veidojas dubultās DNS replikācijas laikā, kā arī savienot divus komplementārus nukleīnskābes fragmentus.
No otras puses, sintāze ir enzīms, kas bioķīmijā katalizē sintēzes procesu. Saskaņā ar EK numuru kategorizāciju tās ir iekļautas liāžu kategorijā.
Nomenklatūra
Atcerieties, ka bioloģiskajā nomenklatūrā sākotnēji tika nošķirtas sintetāzes un sintēzes. Saskaņā ar sākotnējo definīciju sintetāzes kā enerģijas avotu izmanto nukleozīdu trifosfātus (piemēram, ATP, GTP, CTP, TTP un UTP), bet sintēzes to neizmanto.
Tomēr saskaņā ar Apvienotās bioķīmiskās nomenklatūras komisijas (Joint Commission on Biochemical Nomenclature - JCBN) atzinumu terminu "sintēze" var lietot, lai apzīmētu jebkuru fermentu, kas katalizē sintēzi (neatkarīgi no tā, vai tajā tiek izmantoti nukleozīdu trifosfāti), bet terminu "sintēze" drīkst lietot tikai attiecībā uz "ligāzi".
Galvenā atšķirība starp sintēzi un sintēzi ir tāda, ka sintēze ir enzīmu grupa, kas var radīt saites starp molekulām, bet sintēze ir enzīms.
| Sintāzes | Sintetāze |
| Katalizē sintēzes procesu bez ATP | Nepieciešams ATP |
| Ietilpst vāzes vai transferāzes klasifikācijā | Ietilpst ligāžu klasifikācijā |
| piemēram, HMG-COA sintēzi, ATP sintēzi. | piem., sukcīnija1-COA sintetāze, glutamīns sintetāze |
Sintēze vs sintēze Salīdzināšanas tabula
Noskatieties šo videoklipu, lai uzzinātu vairāk par sintēzi un sintēzi
Secinājums
- Sintetāzes darbībai nav nepieciešami NTP, jo tās katalizē sintēzes procesus, kuros nepieciešama nukleozīdu trifosfātu (nevis tikai ATP) hidrolīze.
- 20. gadsimta 80. gados Starptautiskās Bioķīmijas savienības Nomenklatūras komiteja mainīja sintēzes definīciju, iekļaujot tajā visus sintētiskos fermentus neatkarīgi no tā, vai tie izmanto NTP, un sintēze kļuva par ligāzes sinonīmu.
- Ligāze ir enzīms, kas savieno divas mazākas molekulas, izmantojot NTP hidrolīzes enerģiju (parasti kondensācijas reakcijā).