Синтаза мен синтетазаның айырмашылығы неде? (Ашық фактілер) – Барлық айырмашылықтар

Мазмұны
Лигаза ферменттерінің E.C. 6 класына синтазалар мен синтетазалар жатады. Олар синтетикалық реакцияларға қатысады және екі молекуланың бірігуін катализдейді және бір мезгілде АТФ немесе басқа салыстырмалы трифосфаттағы дифосфатты байланысын бұзады.
Синтез кезінде АТФ-ты энергия көзі ретінде пайдаланатын синтетазалардан айырмашылығы биологиялық қосылыстардың, синтазалар - бұл АТФ-ны энергия көзі ретінде пайдаланбай, биологиялық қосылыстардың синтезін катализдейтін кез келген лигазалар.
Бұл мақалада сіз Синтаза мен Синтетазаның айырмашылығын нақты білесіз.
Синтаза дегеніміз не?
Синтаза – биохимиядағы синтез процесін катализдейтін фермент. Биологиялық номенклатура бастапқыда синтетазалар мен синтазаларды ажырататынын есте сақтаңыз.
Бастапқы анықтамаға сәйкес синтетазалар энергия көзі ретінде нуклеозидтрифосфаттарды (мысалы, ATP, GTP, CTP, TTP және UTP) пайдаланады, ал синтазалар қолданбайды.
Дегенмен, биохимиялық номенклатура бойынша Біріккен комиссияның (JCBN) мәліметтері бойынша, «синтаза» синтезді катализдейтін кез келген ферментке сілтеме жасау үшін қолданылуы мүмкін (онда нуклеозидтрифосфаттарды пайдаланатынына қарамастан), бірақ «синтетаза» ” тек “лигазаға” сілтеме жасау үшін қолданылуы керек.
Міне, синтазаның әртүрлі түрлерінің мысалдарының тізімі:
- АТФ синтаза
- Цитрат синтаза
- Триптофансинтезді катализдейтін кез келген ферментке сілтеме жасау үшін қолданылуы мүмкін (онда нуклеозидтрифосфаттарды пайдаланатынына қарамастан), бірақ «синтетаза» тек «лигазаға» сілтеме жасау үшін қолданылуы керек.
Синтаза мен Синтетаза арасындағы негізгі айырмашылық мынада: Синтетаза - молекулалар арасында байланыс түзе алатын ферменттер тобы, ал Синтаза - фермент.
Синтаза Синтетаза АТФ жоқ синтетикалық процесті катализдейді АТФ қажет Ваза немесе трансфераза классификациясына сәйкес келеді Лигаза классификациясына жатады мыс. HMG-COA синтаза, ATP синтаза мыс. Succiny1-COA синтетаза, глютамин синтетаза
Синтаза мен синтетазаны салыстыру кестесі
Синтаза мен синтетаза туралы білу үшін осы бейнені қараңыз
Қорытынды
- Синтетазалардың жұмыс істеуі үшін NTP қажет емес, өйткені олар нуклеозид-трифосфаттардың (жай АТФ емес) гидролизін талап ететін синтетикалық процестерді катализдеді.
- Халықаралық одақ. Биохимияның номенклатуралық комитеті 1980 жылдары синтаза анықтамасын NTP пайдаланғанына қарамастан, барлық синтетикалық ферменттерді қамту үшін өзгертті және синтетаза лигазаның синонимі болды.
- Лигаза - екі кіші молекуланы біріктіретін фермент. энергия NTP гидролизінен (әдетте конденсация арқылы).реакция).
- Псевдоуридинсинтаза
- Май қышқылы синтаза
- Целлюлоза синтаза (UDP түзетін)
- Целлюлоза синтаза (ЖІӨ түзетін)
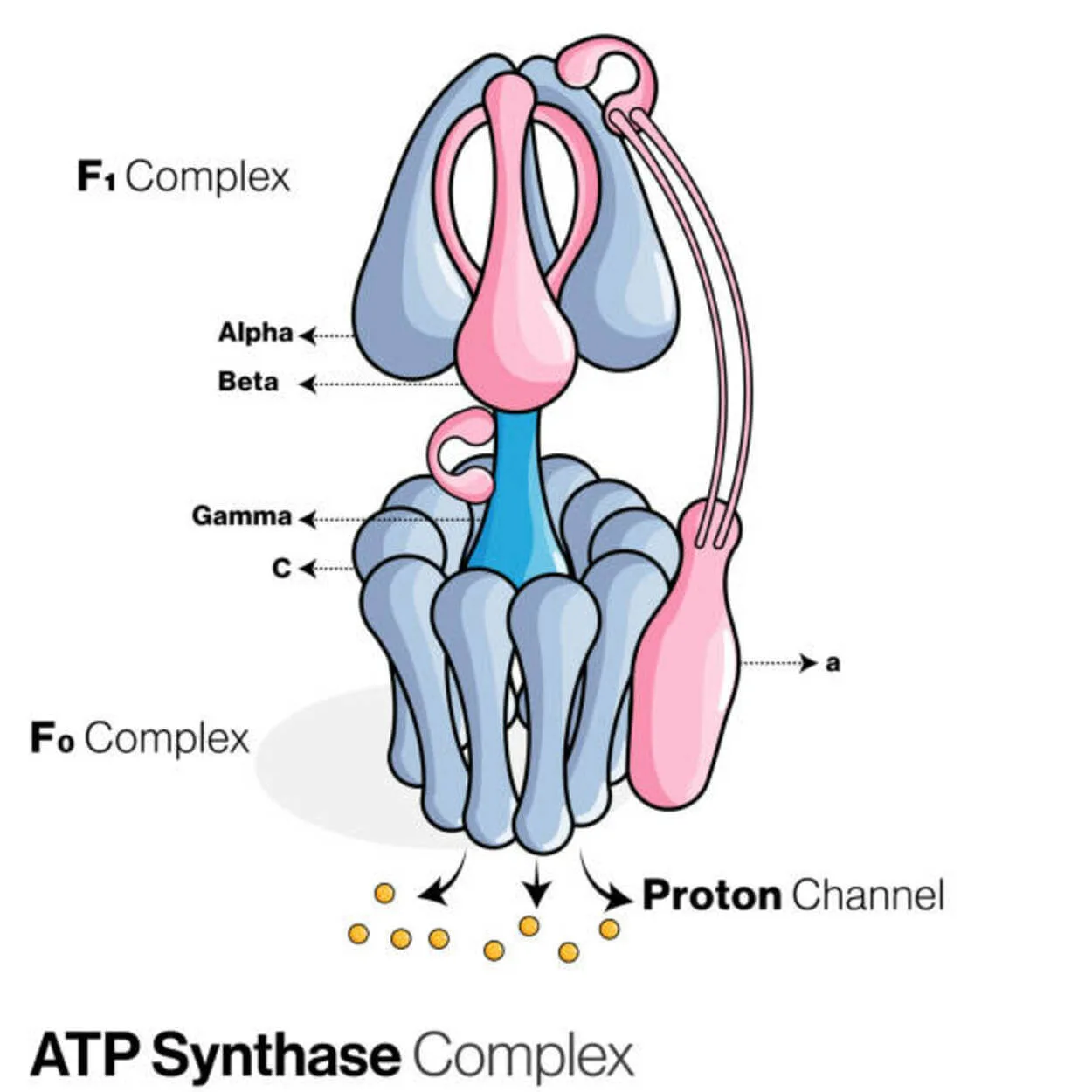
ATP синтаза
Аденозин дифосфаты (АДФ) және бейорганикалық фосфат АТФ синтаза (Pi) деп аталатын ақуыз арқылы аденозинтрифосфат (АТФ) энергия сақтау молекуласын жасау үшін қолданылады.
Ол лигаза ретінде жіктеледі, өйткені ол P-O байланысын (фосфодиэфирлік байланыс) құру арқылы ADP-ны өзгертеді. ATP синтаза деп аталатын молекулалық құрылғы.
Энергия тұрғысынан ADP және Pi-ден ATP өндірісі қажет емес және процесс әдетте басқа жолмен жүреді.
Сондай-ақ_қараңыз: Пибальд жамылғылы хамелеон мен перделенген хамелеон (зерттелді) арасындағы айырмашылық неде - барлық айырмашылықтарЭукариоттардың ішкі митохондриялық мембранасы немесе бактериялардағы плазмалық мембрана арқылы өтетін протон (H+) концентрациясының градиенті жасушалық тыныс алу кезінде АТФ синтезін градиентке қосу арқылы бұл реакцияны алға жылжытады.
Өсімдіктерде АТФ синтаза фотосинтез кезінде АТФ өндіру үшін тилакоидты люменде тилакоидты мембрана арқылы және хлоропласт стромасында түзілетін протон градиентін пайдаланады.
АТФаза үшін эукариоттық АТФ синтазалары F болып табылады. - «кері» функцияны ATPas. Бұл түр бірінші кезекте осы мақалада талқыланады. F-ATPase FO және F1 бөлімшелерінің ATP синтезін қамтамасыз ететін айналмалы қозғалтқыш механизмі бар.
Синтазаның әр түрлі түрлері бар
Цитрат Синтаза
Барлық дерлік тірі жасушаларда цитрат синтаза ферменті бар,лимон қышқылы циклінің бірінші қадамында кардиостимулятор ретінде қызмет етеді және EC 2.3.3.1 (бұрын 4.1.3.7) деп белгіленген. (немесе Кребс циклі).
Цитратсинтаза эукариоттық жасушалардың митохондриялық матрицасында орналасқан, дегенмен оны митохондриялық ДНҚ емес, ядролық ДНҚ кодтайды.
Ол цитоплазмада цитоплазмалық рибосомалар арқылы жасалады және кейіннен митохондриялық матрицаға ауысады.
Бұзылған митохондриялардың болуы үшін типтік сандық фермент маркері цитратсинтаза болып табылады. Цитрат синтазасының ең жоғары белсенділігі қаңқа бұлшықетінде қанша митохондрия бар екенін көрсетеді.
Жоғары интенсивті аралық жаттығулар шыдамдылық жаттығуларына немесе жоғары қарқынды интервал жаттығуларына қарағанда максималды белсенділікті арттыру мүмкіндігіне ие.
Ацетил коферментінде екі көміртегі ацетаты қалдығы және төрт молекула бар. -көміртегі оксалоацетат конденсацияланып, алты көміртекті цитрат түзеді, ол цитратсинтаза катализдейтін конденсация реакциясы нәтижесінде түзіледі.
Триптофансинтаза
Триптофан өндірісінің соңғы екі сатысы: триптофансинтетаза деп те белгілі триптофансинтаза ферментімен катализденеді.
Эубактериялар, архебактериялар, протисталар, саңырауқұлақтар және планталар оның жиі қожайыны болып табылады. Алайда Animalia-да ол жоқ. Әдетте, ол 2 2 тетрамер ретінде көрінеді.
Субірліктер индол-3-глицерин фосфатының қайтымды айналуын катализдейді.индол және глицеральдегид-3-фосфат (G3P) (IGP).
Пиридоксальфосфатқа (ПЛП) тәуелді процесте суббірліктер триптофан түзу үшін индол мен сериннің қайтымсыз конденсациясын катализдейді.
Ұзындығы 25 ангстремді құрайтын және ферментте орналасқан ішкі гидрофобты арна әрбір белсенді аймақты көршілес белсенді аймақпен байланыстырады.
Бұл субстраттың арналануына ықпал етеді, бұл белсенді жерлерде индол түзілетін механизм. басқа белсенді жерлерге тікелей таралады. Триптофан синтазасының құрамында аллостериялық байланысқан белсенді учаскелер бар.
Эубактериялар, архебактериялар, протисталар, саңырауқұлақтар және өсімдіктер триптофан синтазасын қосатыны жиі кездеседі. Адамдарға және басқа жануарларға бұл жетіспейді.
Адамға қажетті тоғыз аминқышқылының бірі, триптофан жиырма стандартты аминқышқылдарының бірі болып табылады. Сондықтан триптофан адам диетасы үшін өте маңызды.
Сондай-ақ, триптофан синтетазасы триптофанның баламалы аналогтарын өндіру үшін субстраттар ретінде фторланған немесе метилденген индолдар сияқты индол аналогтарын пайдалана алатыны белгілі.
Псевдуридин
Грек әрпі psi- псевдоуридинді қысқарту үшін қолданылады, урацил көміртек атомымен азот-көміртек гликозидтік байланысымен емес, көміртегі-көміртекті байланыс арқылы қосылған нуклеозид-уридиннің изомері. (Урацил кейде бұл құрылымда «псевдоурацил» деп аталады.)
Ең таралған РНҚЖасушалық РНҚ-ның өзгеруі псевдоуридин болып табылады. РНҚ транскрипция және синтез кезінде 100-ден астам химиялық бірегей өзгерістерге ұшырауы мүмкін.
Төрт кәдімгі нуклеотидтерге қосымша, бұлар транскрипциядан кейінгі РНҚ экспрессиясына әлеуетті әсер етуі мүмкін және жасушада РНҚ трансляциясы, локализациясы және тұрақтылығы сияқты бірқатар функцияларға ие болуы мүмкін.
Олардың бірі псевдоуридин, уридиндегі C1-N1 типтік байланысының орнына рибоза қантының C1 мен урацилдің С5 арасында С-С байланысы бар уридиннің С5-гликозидті изомері.
С-С байланысына байланысты оның қосымша айналмалы қозғалғыштығы мен конформациялық икемділігі бар. Сонымен қатар, псевдоуридиннің N1 позициясы қосымша сутегі байланысының донорына ие.
Псевдуридин, сондай-ақ 5-рибосилурацил деп те аталады, құрылымдық РНҚ-ның (трансфер, рибосомалық, кіші ядролық (snRNA) және) таныс, бірақ жұмбақ компоненті. шағын ядролық). Ол жақында РНҚ кодтауында да табылды.
Ол бірінші рет анықталған, ең көп таралған және өмірдің барлық үш эволюциялық аймағында кездеседі. Ашытқы тРНҚ-да псевдоуридин нуклеотидтердің шамамен 4%-ын құрайды
Сондай-ақ_қараңыз: Жергілікті диск C және D (толық түсіндірілді) – барлық айырмашылықтарСумен қосымша сутектік байланыстардың түзілуі арқылы бұл негіздің өзгеруі РНҚ-ны тұрақтандыруға және негізді жинақтауды күшейтуге қабілетті.
псевдуридиндердің саны организмнің күрделілігіне қарай өседі. Құрамында 11 псевдоуридин барішек таяқшасының рРНҚ-сы, ашытқылардың цитоплазмалық рРНҚ-да 30, митохондриялық 21S рРНҚ-да бір өзгеріс және адамның рРНҚ-да шамамен 100.
Псевдуридиннің rNARNA- және жұқа түнде түзілетіні көрсетілді. және аймақтық құрылымды тұрақтандырады және олардың мРНҚ декодтауындағы, рибосомаларды құрастырудағы, өңдеудегі және трансляциясындағы рөлдерін қолдауға көмектеседі.
СнРНҚ-дағы псевдоуридин сплайсингті реттеуге көмектесу үшін пре-мРНҚ мен сплайцеосомалық РНҚ арасындағы интерфейсті жақсартатыны көрсетілді.
Май қышқылы синтазасы
FASN Адамдардағы ген май қышқылы синтазасы (FAS) деп аталатын ферментті кодтайды. Май қышқылы синтазасы деп аталатын көп ферментті ақуыз май қышқылдарының синтезін катализдейді.
Бұл екі бірдей 272 кДа көп функционалды полипептидтерден тұратын, бір функционалды аймақтан келесісіне субстраттарды тасымалдайтын бір фермент емес, тұтас ферменттік жүйе.
Оның негізгі жұмысы ацетил- және малонил-КоА
Ацетил-КоА және малонилден пальмитатты (C16:0, ұзын тізбекті қаныққан май қышқылы) түзуді катализдеу үшін NADPH қолдану болып табылады. -КоА декарбоксилдеуші Клейзен конденсациясының реттілігі арқылы май қышқылдарына айналады.
Ұзартудың әрбір айналымынан кейін кеторедуктаза (KR), дегидратаза (DH) және энилредуктаза бета-кето тобын толығымен қаныққан көміртегі тізбегіне дейін азайту үшін ретпен жұмыс істейді.(ER).
Май қышқылдарының тізбегі ұзындығы 16 көміртекке жеткенде, ол ацил тасымалдаушы ақуыздың (ACP) фосфопантетейн протездік тобымен ковалентті байланысқан тиоэстеразаның (ТЭ) әсерінен бөлінеді. (пальмитин қышқылы).
Целлюлоза синтазасы (UDP-түзетін)
Целлюлозаны өндіруге жауапты негізгі фермент УДФ түзетін түрінде целлюлоза синтаза (EC 2.4.1.12) болып табылады. Ол әдетте UDP-глюкоза деп аталады: (1→4) D-глюканға арналған энзимологияның 4-D-глюкозилтрансферазасы.
GDP-глюкозаны целлюлоза синтазасы (GDP-) деп аталатын байланысты фермент пайдаланады. қалыптастыру) (EC 2.4.1.29). Бактерияларда да, өсімдіктерде де ферменттердің осы отбасының өкілдері болады.
Бактерия мүшелері BcsA (бактериялық целлюлоза синтазасы) немесе CelA ретінде де белгілі болуы мүмкін, ал өсімдік мүшелері әдетте CesA (целлюлоза синтазасы) немесе алыпсатарлық CslA (целлюлоза синтаза тәрізді) (жай «целлюлоза») ретінде белгілі. .
ЦесА өсімдіктерге хлоропласт пайда болған эндосимбиоз нәтижесінде алынған. Глюкозилтрансферазалардың 2-ші тұқымдасына осы (GT2) кіреді.
Жер бетіндегі биомассаның көп бөлігі гликозилтрансферазалар деп аталатын ферменттер арқылы биосинтез және гидролиз арқылы түзіледі.
CesA суперсемьясының құрамында болатыны белгілі. жеті топша, ал біріктірілген өсімдік-балдыр супертұқымдасында 10 бар.
Бұл ферментке ие жалғыз жануар тобыурохордаттар, олар оны гендердің көлденең трансферті арқылы 530 миллионнан астам жыл бұрын алған.
Целлюлоза синтазасы (ЖІӨ түзетін)
Бұл фермент гликозилтрансферазалардың гексосильтрансфераза топшасының мүшесі болып табылады. Бұл фермент класы ғылыми атауымен аталады, GDP-глюкоза:1,4-бета-D-глюкан 4-бета-D-глюкозилтрансфераза.
Жиі қолданылатын басқа атаулар - целлюлоза синтаза (гуанозиндифосфат түзетін), целлюлоза синтетаза және гуанозиндифосфоглюкоза-1,4-бета-глюканглюкозилтрансфераза. Бұл фермент сахароза мен крахмал алмасуына қатысады.
Синтетаза дегеніміз не?
Кейде «лигаза» деп аталатын «синтетаза» термині химиялық энергияны үнемдейтін реакцияларды катализдейтін және энергияны тұтынатын ыдырау оқиғалары мен өнімділік арасында делдал болатын шамамен 50 ферменттер класының кез келгеніне қатысты. синтетикалық процестер.
Энергетикалық фосфаттық байланысты ыдырату арқылы олар екі молекуланың қосылуын катализдеу үшін қажетті энергияны тудырады (көп жағдайда аденозинтрифосфатты [АТФ] аденозин дифосфатқа [АДФ] бір мезгілде түрлендіру арқылы) .
Амин қышқылы-РНҚ лигазасы ретінде белгілі лигаза тасымалдаушы РНҚ мен амин қышқылы арасында көміртегі-оттегі байланысының түзілуін катализдейді.
Кейбір ферменттер, мысалы, амидтік синтетазалар және пептидтік синтетазалар белсенді болғанда, көміртегі-азот (C-N)байланыстар түзіледі.
Синтетаза лигаза деп те аталады
Синтетаза мен синтаза арасындағы айырмашылық
Синтетаза - бұл қосылыстардың қосылуын катализдейтін фермент. екі үлкен молекула жаңа химиялық байланысты құру арқылы, әдетте үлкенірек молекулалардың бірінде кішігірім химиялық топтың бір мезгілде гидролизімен немесе ол екі қосылыстардың қосылуын катализдей алады, мысалы, СО, CS, C-N және т.б.
Лигаза әдетте келесі реакцияны тудырады:
- A-C + b = Ab + C
- A+D + B + C + D + E + F = Ab + cD
Тәуелді, шағын топтар кіші әріптермен берілген. Лигаза репликация кезінде қос тізбекті ДНҚ-да дамитын бір тізбекті үзілістерді қалпына келтіре алады, сондай-ақ екі комплементарлы нуклеин қышқылы фрагменттерін байланыстырады.
Екінші жағынан, синтаза биохимиядағы синтез процесін катализдейтін фермент болып табылады. Олар EC санының категориясына сәйкес лиазалар санатына кіреді.
Номенклатура
Биологиялық номенклатура бастапқыда синтетаза мен синтазаны ажырататынын есте сақтаңыз. Бастапқы анықтамаға сәйкес синтетазалар энергия көзі ретінде нуклеозидтрифосфаттарды (мысалы, ATP, GTP, CTP, TTP және UTP) пайдаланады, ал синтазалар қолданбайды.
Соған қарамастан, биохимиялық номенклатура бойынша бірлескен комиссияның (JCBN) мәліметтері бойынша, «синтаза»