Mis on erinevus süntaasi ja süntesaasi vahel? (Faktid selgunud) - kõik erinevused

Sisukord
E.C. 6 ligaasi ensüümide klassi kuuluvad süntaasid ja süntaasid. Nad osalevad sünteetilistes reaktsioonides ja katalüüsivad kahe molekuli ühendamist, lõhkudes samal ajal ATP või muu võrreldava trifosfaadi difosfaatühendust.
Erinevalt süntaasidest, mis kasutavad bioloogiliste ühendite sünteesi käigus ATP-d energiaallikana, on süntaasid mis tahes ligaasid, mis katalüüsivad bioloogiliste ühendite sünteesi ilma ATP-d energiaallikana kasutamata.
Selles artiklis saate te täpselt teada, mis vahe on süntaasi ja süntesaasi vahel.
Mis on süntaas?
Süntaas on ensüüm, mis katalüüsib sünteesiprotsessi biokeemias. Pidage meeles, et bioloogilises nomenklatuuris eristati algselt süntaase ja süntaase.
Vastavalt algsele määratlusele kasutavad süntaasid energiaallikana nukleosiidtrifosfaate (nagu ATP, GTP, CTP, TTP ja UTP), süntaasid aga mitte.
Sellest hoolimata võib vastavalt Joint Commission on Biochemical Nomenclature (JCBN) kohaselt kasutada sõna "süntaas" mis tahes ensüümi kohta, mis katalüüsib sünteesi (sõltumata sellest, kas see kasutab nukleosiidtrifosfaate), kuid sõna "süntaas" tuleks kasutada ainult "ligaasi" kohta.
Siin on loetelu erinevat tüüpi süntaaside näidetest:
- ATP süntaas
- Tsitraadi süntaas
- Trüptofaani süntaas
- Pseudouridiini süntaas
- Rasvhappe süntaas
- Tselluloosisüntaas (UDP moodustav)
- Tselluloosisüntaas (GDP-d moodustav)
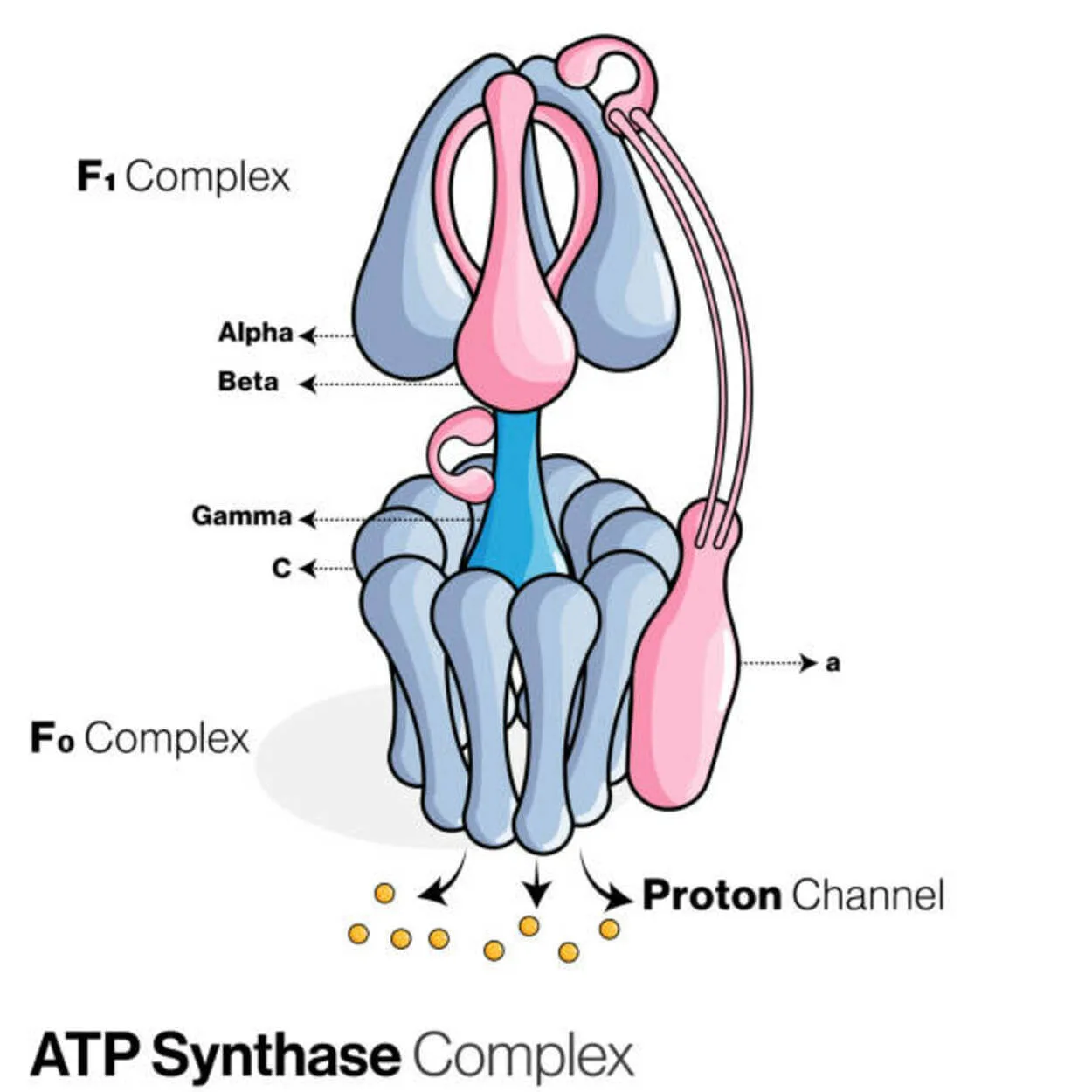
ATP süntaas
Adenosiindifosfaati (ADP) ja anorgaanilist fosfaati kasutab ATP-süntaasi (Pi) nime kandev valk, et luua energiasalvestusmolekul adenosiintrifosfaat (ATP).
Seda liigitatakse ligaasiks, kuna see modifitseerib ADP-d, moodustades P-O sideme (fosfodiester sideme). Molekulaarse seadme nimega ATP-süntaas.
Energia seisukohast on ATP tootmine ADP-st ja Pi-st ebasoovitav ja protsess kulgeb tavaliselt vastupidi.
Prootonite (H+) kontsentratsioonigradient läbi sisemise mitokondri membraani eukarüootides või plasmamembraani bakterites viib seda reaktsiooni edasi, sidudes ATP sünteesi raku hingamise ajal gradienti.
Taimedes kasutab ATP-süntaas fotosünteesi ajal ATP tootmiseks protoonide gradienti, mis moodustub tülakoidi luumenis üle tülakoidimembraani ja kloroplastide stroomasse.
ATPaasi puhul on eukarüootilised ATP-süntaasid F-ATPaasid, mis toimivad "tagurpidi". Seda liiki käsitletakse peamiselt käesolevas artiklis. F-ATPaasi FO ja F1 allühikutel on pöörlemismootori mehhanism, mis võimaldab ATP sünteesi.
On olemas erinevaid süntaasi tüüpe
Tsitraadi süntaas
Peaaegu kõik elusrakud sisaldavad ensüümi tsitraadisüntaasi, mis toimib sammuandjana sidrunhappetsükli esimeses etapis ja mida nimetatakse E.C. 2.3.3.1 (varem 4.1.3.7). (või Krebs'i tsükkel).
Tsitraadisüntaas asub eukarüootiliste rakkude mitokondriaalmaatriksis, kuigi seda kodeerib tuuma-DNA, mitte mitokondriaalne DNA.
See luuakse tsütoplasmas tsütoplasma ribosoomide poolt ja viiakse seejärel mitokondriumi maatriksisse.
Intaktsete mitokondrite olemasolu tüüpiline kvantitatiivne ensüümimarker on tsitraadisüntaas. Tsitraadisüntaasi tippaktiivsus näitab, kui palju mitokondreid on skeletilihases olemas.
Kõrge intensiivsusega intervalltreening võib tõsta maksimaalset aktiivsust rohkem kui vastupidavustreening või kõrge intensiivsusega intervalltreening.
Atsetüülkoensüüm A sisaldab kahesüsivesinikulist atsetaatjääki ja neljasüsivesinikulist oksaloatsetaadi molekuli kondenseerub kuuesüsivesinikulist tsitraati, mis tekib tsitraadisüntaasi poolt katalüüsitud kondensatsioonireaktsioonis.
Trüptofaani süntaas
Kaks viimast sammu trüptofaani tootmisel katalüüsib ensüüm trüptofaanisüntaas, mida nimetatakse ka trüptofaanisüntaasiks.
Eubakterid, Archaebacteria, Protista, Fungi ja Plantae on selle sagedased peremehed. Animalia aga ei oma seda. Tavaliselt esineb see 2 2 tetramerina.
Alamühikud katalüüsivad indool-3-glütseroolfosfaadi pöörduvat muundamist indooliks ja glütseraldehüüd-3-fosfaadiks (G3P) (IGP).
Püridoksaalfosfaadist (PLP) sõltuvas protsessis katalüüsivad allüksused indooli ja seriini pöördumatut kondenseerimist, et tekitada trüptofaani.
Ensüümis asuv 25 angströmi pikkune sisemine hüdrofoobne kanal ühendab iga aktiivse koha naaberaktiivse kohaga.
See soodustab substraadi kanaliseerimist, mis on mehhanism, mille abil aktiivsetes kohtades toodetud indool difundeerub otse teistesse aktiivsetesse kohtadesse. Triptofaanisüntaas sisaldab allostriliselt seotud aktiivseid kohti.
Eubakterid, arheebakterid, protista, seened ja plantae sisaldavad sageli trüptofaanisüntaasi. Inimesel ja teistel loomadel see puudub.
Triptofaan on üks üheksast inimesele vajalikust aminohappest ja kuulub kahekümne standardse aminohappe hulka. Seega on triptofaan inimese toitumises hädavajalik.
Samuti on teada, et trüptofaani süntesaas võib kasutada indoolanalooge, näiteks fluoritud või metüülitud indoole, substraatidena, et toota samaväärseid trüptofaani analooge.
Pseudouridiin
Kreeka tähte psi- kasutatakse pseudouridiini lühendamiseks, mis on nukleosiidi uridiini isomeer, milles uratsiil on süsinikuaatomiga ühendatud süsinik-süsinikühenduse, mitte lämmastik-süsinik glükosiidse sideme kaudu (uratsiili nimetatakse mõnikord "pseudouratsiiliks" sellises paigutuses).
Vaata ka: Erinevus 5.56 ja 22LR vahel (selgitatud!) - kõik erinevusedKõige levinum RNA muutus raku RNAs on pseudouridiin. RNA võib transkriptsiooni ja sünteesi käigus läbida üle 100 keemiliselt unikaalse muutuse.
Lisaks neljale tavapärasele nukleotiidile võivad need potentsiaalselt mõjutada RNA ekspressiooni transkriptsioonijärgselt ja neil on rakus mitmeid funktsioone, sealhulgas RNA translatsioon, lokaliseerimine ja stabiilsus.
Üks neist on pseudouridiin, uridiini C5-glükosiidi isomeer, millel on C-C side riboosisuhkru C1 ja uratsiili C5 vahel uridiinile omase tüüpilise C1-N1 sideme asemel.
Sellel on C-C sideme tõttu täiendav pöörlemisliikuvus ja konformatsiooniline paindlikkus. Lisaks sellele on pseudouridiini N1-positsioonis täiendav vesiniksidemete doonor.
Pseudouridiin, mida nimetatakse ka 5-ribosüüluratsiiliks, on tuttav, kuid salapärane komponent struktuurilistes RNA-des (ülekande-, ribosomaalne, väike tuuma- (snRNA) ja väike nukleolaarne). Hiljuti leiti see ka kodeerivates RNA-des.
See oli esimene, mis avastati, on kõige levinum ja seda võib leida kõigis kolmes evolutsioonilises eluvaldkonnas. Pärmi tRNA-s moodustab pseudouridiin umbes 4% nukleotiididest.
Täiendavate vesiniksidemete moodustamise kaudu veega suudab see alusmuudatus stabiliseerida RNA-d ja suurendada baasihunnikut.
Pseudouridiinide arv kasvab koos organismi keerukusega. 11 pseudouridiini on Escherichia coli rRNA-s, 30 pärmi tsütoplasma rRNA-s, üks muudatus mitokondriaalse 21S rRNA-s ja ligikaudu 100 inimese rRNA-s.
On näidatud, et pseudouridiin rRNAs ja tRNAs peenhäälestab ja stabiliseerib piirkondlikku struktuuri ning aitab säilitada nende rolli mRNA dekodeerimisel, ribosoomi kokkupanekul, töötlemisel ja translatsioonil.
On näidatud, et pseudouridiin snRNAs parandab pre-mRNA ja spliksosomaalse RNA vahelist liideseid, et aidata splaissimist reguleerida.
Rasvhapete süntaas
Inimese FASN geen kodeerib ensüümi, mida nimetatakse rasvhapete süntaasiks (FAS). Mitme ensüümiga valk, mida nimetatakse rasvhapete süntaasiks, katalüüsib rasvhapete sünteesi.
See on terve ensüümsüsteem, mitte ainult üks ensüüm, mis koosneb kahest identsest 272 kDa suurusest multifunktsionaalsest polüpeptiidist, mis edastavad substraate ühest funktsionaalsest domeenist teise.
Selle peamine ülesanne on kasutada NADPH-d, et katalüüsida palmitaadi (C16:0, pikaahelaline küllastunud rasvhape) loomist atsetüül- ja malonüül-CoA-st.
Atsetüül-CoA ja malonüül-CoA muundatakse rasvhapeteks dekarboksüleerivate Claiseni kondensatsiooniprotsesside kaudu.
Pärast iga pikendamise vooru töötavad ketoreduktaas (KR), dehüdrataas (DH) ja enoüülreduktaas järjestikku, et vähendada beeta-ketorühma täielikult küllastunud süsinikuahelaks (ER).
Kui rasvhappeahel on kasvanud 16 süsiniku pikkuseks, vabaneb see tioesteraasi (TE) toimel, mis on kovalentselt seotud atsüülkandja valgu (palmitiinhape) fosfopanteteiini proteetilise rühmaga.
Tselluloosisüntaas (UDP moodustamine)
Peamine tselluloosi tootmise eest vastutav ensüüm on tselluloosisüntaas (EC 2.4.1.12) selle UDP-d moodustavas vormis. Seda nimetatakse üldiselt UDP-glükoosiks: (1→4) ensümaatika 4-D-glükosüültransferaasiks D-glükaani jaoks.
GDP-glükoosi kasutab sellega seotud ensüüm, mida nimetatakse tselluloosisüntaasiks (GDP-d moodustav) (EC 2.4.1.29). Nii bakteritel kui ka taimedel on selle ensüümiperekonna liikmeid.
Bakteriaalsed liikmed võivad olla tuntud ka kui BcsA (bakteriaalne tselluloosisüntaas) või CelA, samas kui taimsed liikmed on tavaliselt tuntud kui CesA (tselluloosisüntaas) või spekulatiivne CslA (tselluloosisüntaasilaadne) (lihtsalt "tselluloos").
CesA omandasid taimed endosümbioosi tulemusena, mille tulemusel tekkis kloroplast. 2. perekonna glükosüültransferaaside hulka kuulub see (GT2).
Suurem osa Maa biomassist toodetakse biosünteesi ja hüdrolüüsi teel ensüümide, nn glükosüültransferaaside abil.
Taimede CesA superperekond sisaldab teadaolevalt seitse alamperekonda ja kombineeritud taimede-vetikate superperekond sisaldab 10 alamperekonda.
Ainus loomarühm, kes omab seda ensüümi, on urohordatid, kes said selle horisontaalse geenisiirde teel üle 530 miljoni aasta tagasi.
Tselluloosisüntaas (GDP moodustav)
See ensüüm kuulub glükosüültransferaaside heksosüültransferaaside alamperekonda. Seda ensüümiklassi nimetatakse teadusliku nimega GDP-glükoos:1,4-beeta-D-glükaan 4-beeta-D-glükosüültransferaas.
Teised sageli kasutatavad nimetused on tselluloosisüntaas (guanosiinidifosfaati moodustav), tselluloosisüntaas ja guanosiinidifosfoglükoos-1,4-beeta-glükaan-glükosüültransferaas. See ensüüm osaleb sahharoosi ja tärklise ainevahetuses.
Mis on süntesaas?
Mõiste "süntesaas", mida mõnikord nimetatakse ka "ligaasiks", viitab mis tahes ühele umbes 50 ensüümi klassist, mis katalüüsivad keemilisi energiat säästvaid reaktsioone ja vahendavad energiat tarbivate lagunemissündmuste ja produktiivsete sünteesiprotsesside vahel.
Lõhestades energeetilist fosfaatsidet, tekitavad nad vajaliku energia kahe molekuli ühendamiseks (paljudel juhtudel adenosiintrifosfaadi [ATP] samaaegse muundamise teel adenosiindifosfaadiks [ADP]).
Aminohappe-RNA ligaasina tuntud ligaas katalüüsib süsinik-hapniku sideme loomist transfeer-RNA ja aminohappe vahel.
Kui teatavad ensüümid, näiteks amiidsünteaasid ja peptiidsünteaasid, on aktiivsed, tekivad süsiniku ja lämmastiku (C-N) sidemed.
Süntaasi tuntakse ka kui ligaasi
Süntaasi ja süntaasi erinevus
Süntaas on ensüüm, mis võib katalüüsida kahe suure molekuli ühendamist, luues uue keemilise sideme, tavaliselt ühe suurema molekuli väikese külgneva keemilise rühma samaaegse hüdrolüüsiga, või ta võib katalüüsida kahe ühendi ühendamist, näiteks C-O, C-S, C-N jne. ühendamist.
Ligaas põhjustab tavaliselt järgmise reaktsiooni:
- A-C + b = Ab + C
- A+D + B + C + D + E + F = Ab + cD
Kui sõltuvad, pisikesed rühmitused on kujutatud väiketähtedega. Ligas võib parandada kahesuunalise DNA replikatsiooni käigus tekkivaid üheahelalisi katkestusi, samuti ühendada kaks komplementaarset nukleiinhappefragmenti.
Teisalt on süntaas ensüüm, mis katalüüsib biokeemias sünteesiprotsessi. EÜ numbrite liigituse järgi kuuluvad nad lüaaside kategooriasse.
Nomenklatuur
Pidage meeles, et bioloogilises nomenklatuuris eristati algselt süntaase ja süntaase. Vastavalt algsele määratlusele kasutavad süntaasid energiaallikana nukleosiidtrifosfaate (nagu ATP, GTP, CTP, TTP ja UTP), süntaasid aga mitte.
Sellest hoolimata võib vastavalt Joint Commission on Biochemical Nomenclature (JCBN) kohaselt kasutada sõna "süntaas" mis tahes ensüümi kohta, mis katalüüsib sünteesi (sõltumata sellest, kas see kasutab nukleosiidtrifosfaate), kuid sõna "süntaas" tuleks kasutada ainult "ligaasi" kohta.
Vaata ka: Hiina Hanfu VS Korea Hanbok VS Jaapani Wafuku - kõik erinevusedSüntaasi ja süntaasi peamine erinevus seisneb selles, et süntaas on ensüümide perekond, mis võib luua molekulide vahelisi sidemeid, samas kui süntaas on ensüüm.
| Süntaas | Süntesaas |
| Katalüüsib sünteesiprotsessi ilma ATP-ta | Nõuab ATP-d. |
| Kuulub vase- või transferaasi klassifikatsiooni alla | Kuulub ligaasi klassifikatsiooni alla |
| nt HMG-COA süntaas, ATP süntaas | nt suktsiin1-COA-sünteaas, glutamiin süntesaas |
Süntaasi vs süntaasi võrdlustabel
Vaadake seda videot, et teada saada süntaasi vs süntaasi kohta
Kokkuvõte
- Süntesaasid ei vaja NTP-d toimimiseks, kuna nad katalüüsisid sünteesiprotsesse, mis nõudsid nukleosiidtrifosfaatide (mitte lihtsalt ATP) hüdrolüüsi.
- Rahvusvahelise Biokeemia Liidu nomenklatuurikomitee muutis süntaasi määratlust 1980. aastatel, et hõlmata kõiki sünteetilisi ensüüme, sõltumata sellest, kas nad kasutavad NTP-d või mitte, ja süntaas muutus süntaasi sünonüümiks.
- Ligaas on ensüüm, mis ühendab kaks väiksemat molekuli omavahel, kasutades NTP hüdrolüüsi energiat (tavaliselt kondensatsioonireaktsiooni kaudu).