合成酵素と合成酵素の違いは何ですか?

目次
E.C.6クラスのリガーゼ酵素には合成酵素と合成酵素があり、合成反応に参加し、2つの分子を結合させると同時に、ATPまたはそれに匹敵する他の3リン酸の2リン酸リンクを切断する触媒となる。
生体化合物の合成時にATPをエネルギー源とする合成酵素とは対照的に、ATPをエネルギー源とせずに生体化合物の合成を触媒するあらゆるリガーゼを合成酵素といいます。
今回は、シンターゼとシンセターゼの違いについて、具体的に知っていただきます。
シンターゼとは?
合成酵素とは、生化学の合成過程を触媒する酵素のことで、生物学的命名法では当初、合成酵素と合成酵素を区別していたことに留意してください。
本来の定義では、合成酵素はヌクレオシド三リン酸(ATP、GTP、CTP、TTP、UTPなど)をエネルギー源として利用するが、合成酵素はそうではない。
関連項目: クッパとキングクッパの違い(謎解き) - All The Differencesとはいえ、生化学命名合同委員会(JCBN)によれば、合成を触媒する酵素であれば(ヌクレオシド三リン酸を用いるかどうかにかかわらず)"シンターゼ "は使用できるが、"シンテターゼ "は "リガーゼ "にのみ使用すべきとしている。
ここでは、さまざまな種類の合成酵素の例を紹介します:
- ATPシンターゼ
- クエン酸シンターゼ
- トリプトファンシンターゼ
- プソイドウリジン合成酵素
- 脂肪酸合成酵素(Fatty acid synthase
- セルロース合成酵素(UDP形成)
- セルロース合成酵素(GDP形成)
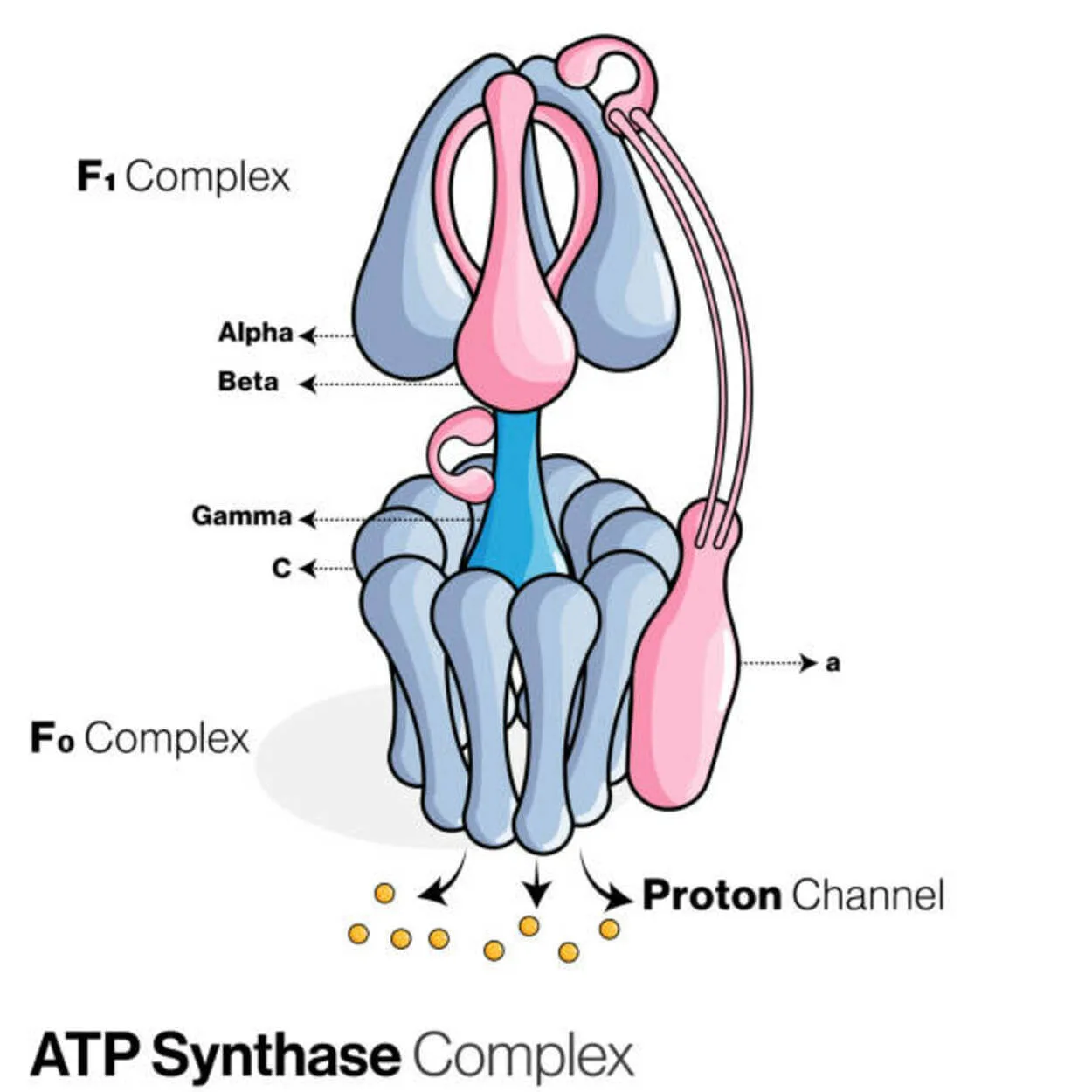
ATPシンターゼ
アデノシン二リン酸(ADP)と無機リン酸は、ATP合成酵素(Pi)というタンパク質によって、エネルギー貯蔵分子であるアデノシン三リン酸(ATP)を作るために使われます。
P-Oリンク(ホスホジエステル結合)を形成してADPを修飾するので、リガーゼに分類される。 ATP合成酵素と呼ばれる分子装置。
エネルギー的には、ADPとPiからATPを生成することは望ましくなく、このプロセスは通常、逆の方向に進むことになる。
真核生物ではミトコンドリア内膜、細菌では細胞膜を横切るプロトン(H+)濃度勾配が、細胞呼吸によるATP合成を勾配に連動させてこの反応を促進する。
植物では、光合成の際に、チラコイド内腔で形成されたプロトン勾配を利用して、チラコイド膜を越えて葉緑体間質に入り、ATPを生成するATP合成酵素。
真核生物のATP合成酵素は、"逆 "に機能するF-ATPaseであり、今回は主にこの種類を取り上げます。 F-ATPaseのFOとF1サブユニットは、ATP合成を可能にする回転モーター機構を持っています。
シンターゼの種類は様々です
クエン酸シンターゼ
ほぼすべての生きた細胞には、クエン酸サイクルの第1ステップでペースメーカーとなるクエン酸合成酵素があり、E.C. 2.3.3.1 (旧4.1.3.7)(またはクレブスサイクル)と呼ばれています。
クエン酸合成酵素は真核細胞のミトコンドリアマトリックスに存在するが、ミトコンドリアDNAではなく核DNAがそれをコードしている。
細胞質で細胞質リボソームによって作られ、その後、ミトコンドリアマトリックスに移動します。
ミトコンドリアの存在を示す代表的な定量的酵素マーカーはクエン酸合成酵素で、クエン酸合成酵素のピーク活性から骨格筋のミトコンドリアの数がわかります。
高強度インターバルトレーニングは、持久力トレーニングや高強度インターバルトレーニングのいずれよりも最大活動量を上げる可能性があります。
アセチルコエンザイムAは炭素数2の酢酸残基を持ち、炭素数4のオキサロ酢酸1分子が縮合して炭素数6のクエン酸を生成し、クエン酸合成酵素が触媒となる縮合反応により生成される。
トリプトファンシンターゼ
トリプトファンの生産における最後の2つのステップは、トリプトファン合成酵素とも呼ばれる酵素によって触媒されます。
真正細菌、古細菌、原生生物、菌類、植物界に多く存在するが、動物界には存在しない。 通常は2 2 4量体として現れる。
サブユニットは、インドール-3-グリセロールリン酸をインドールとグリセルアルデヒド-3-リン酸(G3P)(IGP)に可逆的に変換する触媒である。
ピリドキサールリン酸(PLP)依存的なプロセスで、サブユニットはインドールとセリンの不可逆的な縮合を触媒してトリプトファンを生成する。
酵素内にある長さ25オングストロームの内部疎水チャネルが、各活性部位と隣接する活性部位をつないでいます。
トリプトファン合成酵素には、アロステリックに結合した活性部位があり、活性部位で生成されたインドールが他の活性部位に直接拡散する機構である基質チャネリングが促進される。
真正細菌、古細菌、原生生物、菌類、植物界では、トリプトファン合成酵素を含むことが頻繁に発見されている。 人間や他の動物では、これを欠いている。
トリプトファンは、人間に必要な9種類のアミノ酸のひとつで、20種類の標準アミノ酸のひとつです。 したがって、トリプトファンは人間の食生活に欠かせないものです。
また、トリプトファン合成酵素は、フッ素化インドールやメチル化インドールなどのインドール類縁体を基質として、同等のトリプトファン類縁体を生成できることが知られています。
プソイドウリジン
ギリシャ文字のpsi-は、ウラシルが窒素-炭素のグリコシド結合ではなく、炭素-炭素結合で炭素原子に結合しているヌクレオシド・ウリジンの異性体であるpseudouridineを略すのに使われる(この配列でウラシルは「pseudouracil」と呼ばれることもある)。
細胞内RNAに最も多く見られるRNAの変化はプソイドウリジンである。 RNAは転写と合成の過程で100以上の化学的にユニークな変化を遂げることができる。
従来の4つのヌクレオチドに加え、転写後のRNA発現に影響を与える可能性があり、RNAの翻訳、局在、安定性など、細胞内で多くの機能を持つ可能性があります。
ウリジンのC5-グリコシド異性体で、ウリジンのC1-N1結合の代わりにリボース糖のC1とウラシルのC5との間にC-C結合を持つものがその一つである。
また、プソイドウリジンのN1位には水素結合供与体が存在するため、回転運動性と立体配座の自由度が高い。
プソイドウリジンは、5-リボシルウラシルとも呼ばれ、構造RNA(トランスファー、リボソーム、小核(snRNA)、小核球)の成分として知られていますが、最近、コーディングRNAにも含まれていることが判明しました。
酵母のtRNAでは、プソイドウリジンはヌクレオチドの約4%を占めています。
この塩基の変化は、水との水素結合をさらに形成することで、RNAを安定化させ、塩基のスタッキングを促進することができる。
大腸菌のrRNAには11個、酵母の細胞質rRNAには30個、ミトコンドリアの21S rRNAには1個、ヒトのrRNAには約100個の偽ウリジンが存在し、生物の複雑さとともに増加する。
rRNAやtRNAに含まれるプソイドウリジンは、領域構造を微調整して安定化させ、mRNAの解読、リボソームの組み立て、処理、翻訳における役割の維持を助けることが証明されています。
snRNAに含まれるプソイドウリジンは、プレmRNAとスプライソソームRNAの界面を改善し、スプライシングの制御に役立つことが実証されています。
脂肪酸シンターゼ
ヒトのFASN遺伝子は、脂肪酸合成酵素(FAS)と呼ばれる酵素をコードしています。 脂肪酸合成酵素という複数の酵素を持つタンパク質が、脂肪酸の合成を触媒しています。
それは、1つの酵素だけでなく、酵素システム全体であり、2つの同一の272kDaの多機能ポリペプチドで構成され、基質をある機能ドメインから次の機能ドメインに転送する。
関連項目: F-16 vs F-15- (アメリカ空軍) - All The Differences主な仕事は、NADPHを使って、アセチル-CoAとマロニル-CoAからパルミチン酸(C16:0、長鎖飽和脂肪酸)を作る触媒を行うことである
アセチル-CoAとマロニル-CoAは、一連の脱炭酸型クレーゼン縮合過程を経て脂肪酸に変換される。
伸長のたびに、ケト還元酵素(KR)、脱水酵素(DH)、エノイル還元酵素が順に働き、βケト基が減少して完全飽和炭素鎖(ER)になります。
脂肪酸鎖が炭素数16の長さに成長すると、アシルキャリアプロテイン(ACP)(パルミチン酸)のホスホパンテテインの補欠基と共有結合しているチオエステラーゼ(TE)の働きにより、脂肪酸が放出されます。
セルロース合成酵素(UDP形成)
セルロースを生産する主要な酵素は、UDP形成型のセルロース合成酵素(EC 2.4.1.12)である。 一般に、D-グルカンのUDP-グルコース:(1→4)酵素学の4-D-グルコシルトランスフェラーゼと呼ばれるものである。
GDP-グルコースは、セルロース合成酵素(GDP-forming)(EC 2.4.1.29)という関連酵素によって利用される。 この酵素ファミリーのメンバーは、バクテリアと植物の両方に存在する。
細菌のメンバーはBcsA(細菌性セルロース合成酵素)またはCelAとも呼ばれることがあり、植物のメンバーは通常CesA(セルロース合成酵素)または推測のCslA(セルロース合成酵素様)(単に「セルロース」)と呼ばれています。
CesAは、植物が葉緑体を生み出した内共生によって獲得したものである。 グルコシルトランスフェラーゼのファミリー2には、このもの(GT2)がある。
地球上のバイオマスの大部分は、糖転移酵素と呼ばれる酵素による生合成と加水分解によって生産されています。
植物のCesAスーパーファミリーには7つのサブファミリーがあり、植物と藻類を合わせたスーパーファミリーには10のサブファミリーがあることが知られています。
この酵素を持つ動物群は、5億3,000万年以上前に水平遺伝子移動で手に入れたウロ虫類だけである。
セルロース合成酵素(GDP形成型)
本酵素は、ヘキソシルトランスフェラーゼ・サブファミリーに属し、学名GDP-glucose:1,4-beta-D-glucan 4-beta-D-glucosyltransferase と呼ばれる。
また、セルロース合成酵素(グアノシン二リン酸形成酵素)、セルロース合成酵素、グアノシン二リン酸グルコース-1,4-β-グルカングルコシルトランスフェラーゼという名称もよく使われます。 ショ糖やデンプンの代謝に関与する酵素です。
シンテターゼとは?
化学的なエネルギー保存反応を触媒し、エネルギーを消費する分解過程と生産的な合成過程を仲介する約50種類の酵素のうち、いずれか1つを指す。
エネルギー源であるリン酸結合を切断することで、2つの分子の結合を触媒するのに必要なエネルギーを生成します(多くの場合、アデノシン三リン酸[ATP]とアデノシン二リン酸[ADP]を同時に変換します)。
アミノ酸-RNAリガーゼと呼ばれるリガーゼは、トランスファーRNAとアミノ酸の間に炭素-酸素結合を作ることを触媒するものである。
アミド合成酵素やペプチド合成酵素など、特定の酵素が活性化すると、炭素-窒素(C-N)結合が生成される。
シンセターゼはリガーゼとも呼ばれる
シンテターゼとシンセターゼの違い
合成酵素は、新しい化学結合を作ることによって2つの大きな分子の結合を触媒することができる酵素で、通常、大きな分子の1つ上の小さなペンダント化学基の同時加水分解を伴うか、またはC-O、C-S、C-Nなどの結合など、2つの化合物の連結を触媒することができる酵素です。
リガーゼは通常、以下のような反応を起こします:
- A-C+b=Ab+C
- A+D+B+C+D+E+F=Ab+cD
リガーゼは、複製中の二本鎖DNAに生じた一本鎖切断を修復したり、2つの相補的な核酸断片を連結したりします。
一方、シンターゼは生化学の合成過程を触媒する酵素で、EC番号の分類ではリアーゼのカテゴリーに含まれるそうです。
名称の由来
生物学上の命名法では、当初、ヌクレオシド三リン酸(ATP、GTP、CTP、TTP、UTPなど)をエネルギー源とする合成酵素と、そうでない合成酵素を区別していた。
とはいえ、生化学命名合同委員会(JCBN)によれば、合成を触媒する酵素であれば(ヌクレオシド三リン酸を用いるかどうかにかかわらず)"シンターゼ "は使用できるが、"シンテターゼ "は "リガーゼ "にのみ使用すべきとしている。
シンセターゼとシンテターゼの主な違いは、シンテターゼが分子間の結合を生成する可能性のある酵素の仲間であるのに対し、シンテターゼは酵素の仲間であることです。
| シンターゼ | シンセターゼ |
| ATPがなくても合成過程を触媒する | ATPを必要とする |
| 花器またはトランスフェラーゼの分類に含まれる | リガーゼの分類に含まれる |
| 例:HMG-COAシンターゼ、ATPシンターゼ | 例:Succiny1-COA合成酵素、Glutamine シンテターゼ |
シンセターゼとシンセターゼの比較表
シンセターゼとシンテターゼについて知るには、このビデオをご覧ください。
結論
- 合成酵素は、ヌクレオシド三リン酸(単なるATPではない)の加水分解を必要とする合成過程を触媒するため、機能するためにNTPを必要としません。
- 国際生化学連合の命名法委員会は1980年代に合成酵素の定義を変更し、NTPを利用するか否かにかかわらず、すべての合成酵素を包含するようにし、合成酵素はリガーゼと同義になった。
- リガーゼとは、NTP加水分解によるエネルギーを利用して、2つの小さな分子を結合させる酵素のことです(通常は縮合反応による)。