Synthase와 Synthetase의 차이점은 무엇입니까? (사실 공개) – 모든 차이점

목차
리가제 효소의 E.C. 6 부류에는 신타제 및 신테타제가 포함된다. 그들은 합성 반응에 참여하고 두 분자의 결합을 촉매하는 동시에 ATP 또는 다른 유사한 삼인산에서 이인산 연결을 끊습니다.
합성 과정에서 ATP를 에너지원으로 사용하는 합성 효소와 대조적입니다. 생물학적 화합물 중에서 신타제는 ATP를 에너지원으로 사용하지 않고 생물학적 화합물의 합성을 촉매하는 모든 리가제입니다.
이 기사에서는 Synthase와 Synthetase의 차이점을 정확히 알게 될 것입니다.
또한보십시오: Te와 Tu(스페인어)의 차이점은 무엇입니까? (정교한 관점) – 모든 차이점신타아제란? 합성효소는 생화학에서 합성과정을 촉매하는 효소이다. 생물학적 명명법은 처음에 합성효소와 합성효소를 구분했다는 점을 명심하십시오.
원래의 정의에 따르면 합성효소는 뉴클레오사이드 트리포스페이트(예: ATP, GTP, CTP, TTP 및 UTP)를 에너지원으로 사용하지만 합성효소는 그렇지 않습니다.
그럼에도 불구하고 JCBN(Joint Commission on Biochemical Nomenclature)에 따르면 "합성효소"는 합성을 촉매하는 모든 효소(뉴클레오시드 트리포스페이트를 사용하는지 여부에 관계없이)를 지칭하는 데 사용될 수 있지만 "합성효소"는 "는 "ligase"를 지칭하는 데에만 사용되어야 합니다.
다음은 다양한 유형의 synthase 목록입니다.
- ATP synthase
- 구연산 합성효소
- 트립토판합성을 촉매하는 모든 효소를 지칭하는 데 사용할 수 있지만(뉴클레오사이드 트리포스페이트를 사용하는지 여부에 관계없이) "synthetase"는 "ligase"를 지칭하는 데만 사용해야 합니다. Synthase와 Synthetase의 주요 차이점은 다음과 같습니다. Synthetase는 분자 사이에 결합을 생성할 수 있는 효소군이고 Synthase는 효소입니다.
Synthase 합성효소 ATP 없이 합성 과정을 촉매함 ATP 필요 꽃병 또는 전이효소 분류에 속함 리가아제 분류 에 속함. HMG-COA 합성효소, ATP 합성효소 예. Succiny1-COA synthetase, Glutamine synthetase
Synthase vs Synthetase 비교표
synthase vs synthetase
결론
- 합성효소는 뉴클레오사이드 트리포스페이트(단순히 ATP가 아님)의 가수분해를 필요로 하는 합성 과정을 촉매하기 때문에 NTP가 기능하는 데 필요하지 않습니다.
- International Union of Biochemistry's Nomenclature Committee는 1980년대에 합성 효소의 정의를 변경하여 NTP 사용 여부에 관계없이 모든 합성 효소를 포함하도록 했으며 합성 효소는 리가아제와 동의어가 되었습니다. NTP 가수분해 에너지(일반적으로 응축을 통해)반응).
- 슈도우리딘 합성효소
- 지방산 합성효소
- 셀룰로오스 합성효소(UDP 형성)
- 셀룰로오스 합성효소(GDP 형성)
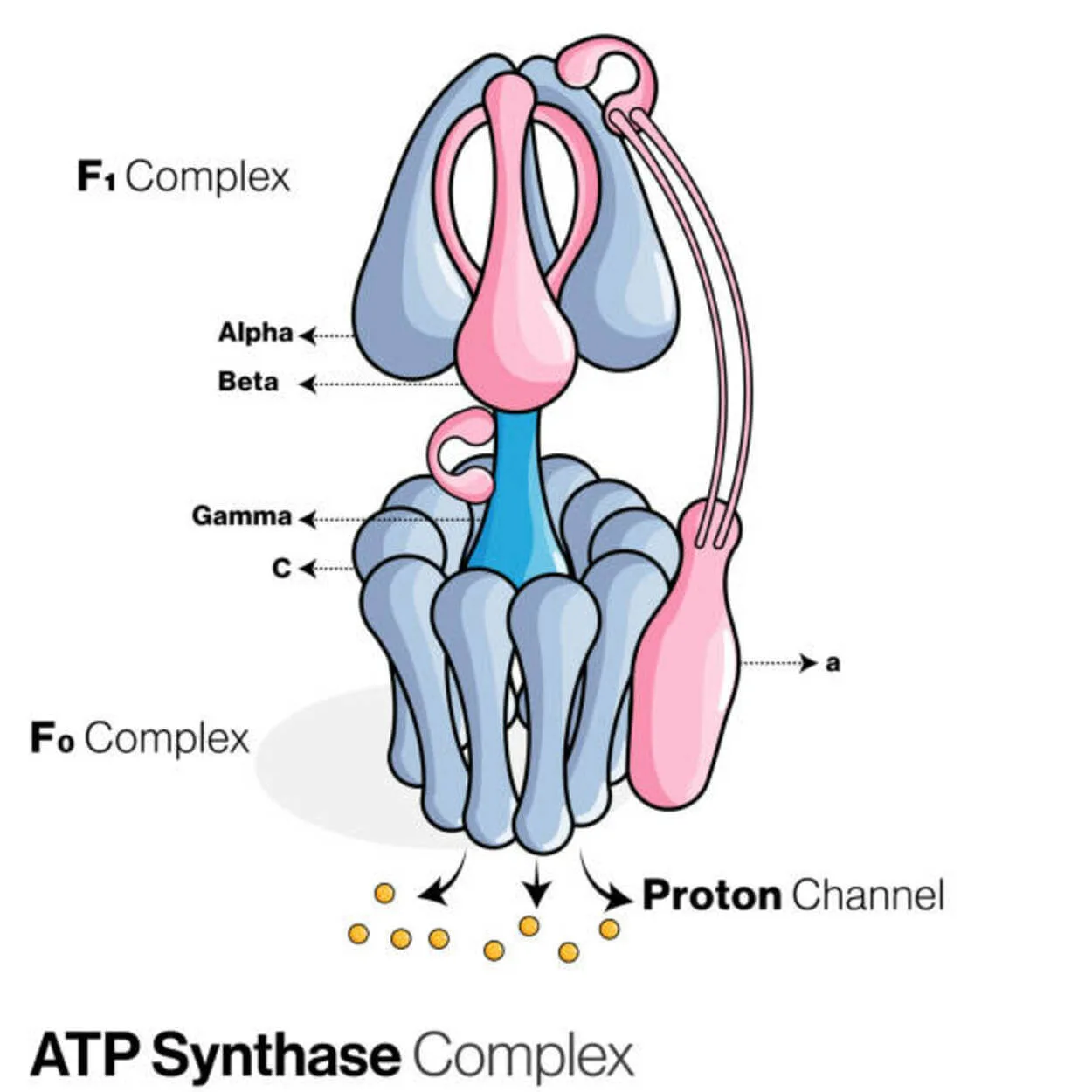
ATP 합성효소
아데노신이인산(ADP)과 무기인산염은 ATP 합성효소(Pi)라는 단백질에 의해 에너지 저장 분자인 아데노신삼인산(ATP)을 생성하는데 사용된다.
P-O 결합(phosphodiester bond)을 형성하여 ADP를 변형시키므로 리가제로 분류된다. ATP 합성 효소라고 하는 분자 장치입니다. 에너지 측면에서 ADP와 Pi에서 ATP를 생산하는 것은 바람직하지 않으며 프로세스는 일반적으로 반대 방향으로 진행됩니다.
진핵생물의 내부 미토콘드리아 막 또는 박테리아의 원형질막을 가로지르는 양성자(H+) 농도 구배는 구배에 대한 세포 호흡 동안 ATP 합성을 결합함으로써 이 반응을 추진합니다. 식물에서 ATP 합성효소는 광합성 동안 ATP를 생성하기 위해 틸라코이드 막을 가로질러 엽록체 기질로 틸라코이드 내강에서 형성된 양성자 구배를 사용합니다.ATPase의 경우 진핵생물 ATP 합성효소는 F -ATPases는 "역방향"으로 작동합니다. 이 유형은 주로 이 문서에서 설명합니다. F-ATPase의 FO 및 F1 서브유닛은 ATP 합성을 가능하게 하는 회전 모터 메커니즘을 가지고 있습니다. 1>거의 모든 살아있는 세포에는 효소 구연산 합성 효소가 포함되어 있습니다.이는 구연산 순환의 첫 번째 단계에서 심박조율기 역할을 하며 EC 2.3.3.1(이전 4.1.3.7)로 지정됩니다. (또는 크렙스 주기).
Citrate synthase는 진핵 세포의 미토콘드리아 기질에 위치하지만 미토콘드리아 DNA가 아닌 핵 DNA에 암호화되어 있습니다.
세포질 리보솜에 의해 세포질에서 생성된 후 미토콘드리아 기질로 이동합니다.
온전한 미토콘드리아의 존재에 대한 전형적인 정량적 효소 마커는 시트레이트 신타아제입니다. Citrate synthase의 최대 활동은 골격근에 얼마나 많은 미토콘드리아가 존재하는지를 보여줍니다.
고강도 인터벌 트레이닝은 지구력 트레이닝이나 고강도 인터벌 트레이닝보다 최대 활동을 높일 수 있는 잠재력이 있습니다.
아세틸 코엔자임 A에는 2개의 탄소 아세테이트 잔기와 4개의 분자가 있습니다. -카본 옥살로아세테이트가 응축되어 6탄소 시트레이트를 생성하며, 이는 시트레이트 합성 효소에 의해 촉매되는 축합 반응에 의해 생성됩니다.
트립토판 합성 효소
트립토판 생산의 마지막 두 단계는 다음과 같습니다. 트립토판 합성 효소라고도 알려진 효소 트립토판 합성 효소에 의해 촉매됩니다.
Eubacteria, Archaebacteria, Protista, Fungi 및 Plantae가 빈번한 숙주입니다. 그러나 Animalia에는 없습니다. 일반적으로 2 2 tetramer로 나타납니다.
서브유닛은 인돌-3-글리세롤 포스페이트에서인돌 및 글리세르알데하이드-3-포스페이트(G3P)(IGP). PLP(pyridoxal phosphate) 의존적 공정에서 서브유닛은 인돌과 세린의 비가역적 축합을 촉매하여 트립토판을 생성합니다. 3>
25 옹스트롬 길이의 내부 소수성 채널은 효소에 위치하여 각 활성 부위를 인접한 활성 부위에 연결합니다. 이는 활성 부위에서 인돌이 생성되는 메커니즘인 기질 채널링을 촉진합니다. 다른 활성 부위로 직접 확산됩니다. 트립토판 신타제는 알로스테릭하게 결합된 활성 부위를 포함합니다. Eubacteria, Archaebacteria, Protista, Fungi 및 Plantae는 트립토판 신타제를 포함하는 것으로 자주 발견됩니다. 인간과 다른 동물들은 그것이 부족합니다.
인간에게 필요한 9가지 필수 아미노산 중 하나인 트립토판은 20가지 표준 아미노산 중 하나입니다. 따라서 트립토판은 인간의 식단에 필수적입니다. 또한 트립토판 합성효소는 플루오르화 또는 메틸화 인돌과 같은 인돌 유사체를 기질로 사용하여 동등한 트립토판 유사체를 생성할 수 있는 것으로 알려져 있습니다.
슈도우리딘
그리스 문자 psi-는 우라실이 질소-탄소 글리코시드 연결이 아닌 탄소-탄소 연결에 의해 탄소 원자에 연결된 뉴클레오시드 우리딘의 이성질체인 슈도우리딘을 약어로 사용합니다. (Uracil은 때때로 이 배열에서 "pseudouracil"로 언급됩니다.)
가장 널리 퍼진 RNA세포 RNA의 변형은 pseudouridine입니다. RNA는 전사 및 합성 동안 100개 이상의 화학적으로 고유한 변경을 겪을 수 있습니다.
4개의 기존 뉴클레오티드 외에도 이들은 잠재적으로 전사 후 RNA 발현에 영향을 미칠 수 있으며 RNA 번역, 국소화 및 안정성을 포함하여 세포에서 여러 기능을 수행합니다. 이들 중 하나는 우리딘에 존재하는 전형적인 C1-N1 결합 대신에 리보스 당의 C1과 우라실의 C5 사이에 C-C 결합을 갖는 우리딘의 C5-글리코시드 이성질체인 슈도우리딘이다.
C-C 결합으로 인해 추가적인 회전 이동성과 구조적 유연성을 갖습니다. 또한 슈도우리딘의 N1 위치는 추가적인 수소 결합 기증자를 보유하고 있습니다. 5-리보실우라실이라고도 하는 슈도우리딘은 구조 RNA(전달, 리보솜, 작은 핵(snRNA) 및 작은 핵소체). 그것은 최근 코딩 RNA에서도 발견되었습니다.
가장 먼저 발견되었고 가장 널리 퍼져 있으며 생명의 세 가지 진화 영역 모두에서 발견될 수 있습니다. 효모 tRNA에서 슈도우리딘은 물과의 추가 수소 결합 형성을 통해 뉴클레오티드의 약 4%를 3>0>만들며, 이 염기 변경은 RNA를 안정화하고 염기 스태킹을 향상시킬 수 있습니다. pseudouridines의 수는 유기체의 복잡성과 함께 증가합니다. 슈도우리딘은 11개대장균의 rRNA, 효모의 세포질 rRNA에 30개, 미토콘드리아의 21S rRNA에 1개, 인간의 rRNA에 대략 100개. 지역 구조를 안정화하고 mRNA 해독, 리보솜 조립, 처리 및 번역에서 역할 유지를 돕습니다.
snRNA의 pseudouridine이 pre-mRNA와 spliceosomal RNA 사이의 계면을 개선하여 스플라이싱을 조절하는 것을 돕는다는 것이 입증되었습니다.
Fatty Acid Synthase
The FASN 인간의 유전자는 지방산 합성 효소(FAS)로 알려진 효소를 암호화합니다. 지방산 합성 효소라고 불리는 다중 효소 단백질은 지방산 합성을 촉매합니다. 기질을 하나의 기능 도메인에서 다음 도메인으로 전달하는 두 개의 동일한 272kDa 다기능 폴리펩타이드로 구성된 하나의 효소가 아닌 전체 효소 시스템입니다.
주요 임무는 NADPH를 사용하여 아세틸- 및 말로닐-CoA
아세틸-CoA 및 말로닐로부터 팔미테이트(C16:0, 장쇄 포화 지방산) 생성을 촉매하는 것입니다. -CoA는 일련의 탈카르복실화 Claisen 응축 과정을 통해 지방산으로 전환됩니다. 3>
연신의 각 라운드에 이어 케토리덕타제(KR), 데히드라타제(DH) 및 에노일 리덕타제가 순차적으로 작용하여 베타 케토기를 완전 포화 탄소 사슬로 환원시킵니다.(응급실).
지방산 사슬이 탄소 16개 길이로 성장하면 아실 운반 단백질(acyl carrier protein, ACP)의 포스포판테테인 보결분자단에 공유결합으로 연결된 티오에스테라아제(thioesterase, TE)의 작용에 의해 지방산 사슬이 방출됩니다. (palmitic acid).
셀룰로오스 합성효소(UDP-형성) 11>
셀룰로오스 생성을 담당하는 주요 효소는 UDP 형성 형태의 셀룰로오스 합성효소(EC 2.4.1.12)입니다. 일반적으로 UDP-glucose라고 한다: (1→4) D-glucan에 대한 Enzymology의 4-D-glucosyltransferase.GDP-glucose는 cellulose synthase(GDP- 성형) (EC 2.4.1.29). 박테리아와 식물은 모두 이 계열의 효소를 가지고 있습니다.
박테리아 구성원은 BcsA(박테리아 셀룰로스 신타제) 또는 CelA로 알려져 있는 반면, 식물 구성원은 일반적으로 CesA(셀룰로스 신타제) 또는 추측성 CslA(셀룰로스 신타제 유사)(간단히 "셀룰로스")로 알려져 있습니다. . 3>
CesA는 엽록체를 발생시키는 세포내 공생의 결과로 식물에 의해 획득되었습니다. 글루코실트랜스퍼라아제의 패밀리 2에는 이 것(GT2)이 포함됩니다.
지구상 바이오매스의 대부분은 글리코실트랜스퍼라아제라고 하는 효소에 의한 생합성 및 가수분해를 통해 생산됩니다.
식물 CesA 슈퍼패밀리는 다음을 포함하는 것으로 알려져 있습니다. 7개의 아과, 결합된 식물-조류 슈퍼패밀리는 10개를 포함합니다.
이 효소를 소유한 유일한 동물 그룹은5억 3천만년 전에 수평적 유전자 전달을 통해 얻은 urochordates.
또한보십시오: 관계와 관계의 차이점 연인 – 모든 차이점셀룰로스 합성 효소(GDP-형성)
이 효소는 글리코실트랜스퍼라제의 헥소실트랜스퍼라제 아과의 구성원입니다. 이 효소 클래스는 과학적 이름인 GDP-글루코스:1,4-베타-D-글루칸 4-베타-D-글루코실트랜스퍼라제로 지칭됩니다. 자주 사용되는 다른 이름은 셀룰로오스 합성효소(구아노신 디포스페이트 형성), 셀룰로오스 합성효소 및 구아노신 디포스포글루코스-1,4-베타-글루칸 글루코실전이효소입니다. 이 효소는 자당과 전분의 대사에 관여합니다.
합성효소란?
"합성효소"(때때로 "리가제"라고도 함)는 화학 에너지 보존 반응을 촉매하고 에너지 소비 분해 사건과 생산적인 반응 사이를 중재하는 대략 50가지 효소 클래스 중 하나를 나타냅니다. 합성 과정. 활성 인산염 결합을 끊음으로써 두 분자의 결합을 촉매하는 데 필요한 에너지를 생성합니다(많은 경우 아데노신 삼인산[ATP]이 아데노신 이인산[ADP]으로 동시에 전환됨) . 3>
아미노산-RNA 리가제로 알려진 리가제는 전달 RNA와 아미노산 사이의 탄소-산소 결합 생성을 촉매하는 것이다.
아마이드 합성효소, 펩타이드 합성효소 등 특정 효소가 활성화되면 탄소-질소(C-N)결합이 생성됩니다.
Synthetase는 Ligase라고도 합니다.
Synthetase와 Synthase의 차이점
Synthetase는 결합을 촉매할 수 있는 효소입니다. 일반적으로 더 큰 분자 중 하나에 있는 작은 펜던트 화학 그룹의 동시 가수분해와 함께 새로운 화학 결합을 생성하여 두 개의 큰 분자를 만들거나 CO, C-S, C-N 등의 결합과 같은 두 화합물의 연결을 촉매할 수 있습니다. .
리가아제는 일반적으로 다음과 같은 반응을 일으킵니다:
- A-C + b = Ab + C
- A+D + B + C + D + E + F = Ab + cD
여기서 종속된 작은 그룹은 소문자로 표시됩니다. 리가아제는 복제 중에 이중 가닥 DNA에서 발생하는 단일 가닥 파손을 복구할 수 있을 뿐만 아니라 두 개의 상보적인 핵산 단편을 연결할 수 있습니다. 한편, 신타아제는 생화학에서 합성 과정을 촉매하는 효소입니다. EC 번호 분류에 따라 분해효소 범주에 포함됩니다.
명명법
생물학적 명명법은 초기에 합성효소와 합성효소를 구별했다는 점을 명심하십시오. 원래의 정의에 따르면 합성효소는 뉴클레오시드 삼인산(예: ATP, GTP, CTP, TTP 및 UTP)을 에너지원으로 활용하지만 합성효소는 그렇지 않습니다.
그럼에도 불구하고 JCBN(Joint Commission on Biochemical Nomenclature)에 따르면 "synthase"